Production and Partial Purification of Thermostable Chitinase from Bacillus cereus BSH-4 and its Antifungal Activity
Basavaraj Shrishailappa Hungund*, Gururaj Basavaraj Tennalli, Sharanappa Achappa, Manisha Nagaraj Divate
Abstract
The study highlights the production of chitinase from Bacillus cereus BSH-4, which was isolated from a marine soil source and identified by 16S rRNA gene sequencing. After 96 hours of incubation in shake flask conditions profiling studies showed maximum enzyme activity and growth. The ammonium sulfate precipitation method was used at 80% saturation for partial enzyme purification with further purification done by dialysis wherein the purification fold increased to 1.75. The sodium dodecyl sulfate gel electrophoresis (SDS-PAGE) and silver staining methods were employed to assess the enzyme molecular weight observed to be 60 kDa. The enzyme activity and stability process parameters effects like temperature and pH were studied. At a pH value of 7.0, the greatest enzyme activity was noted which was relatively retained at 80% till pH 9.0. Similarly, higher enzyme activity was observed at a temperature of 30oC and retained 70% relative activity till 60oC, indicating the thermostable nature of the enzyme. Studies on the influence of metal ions showed that Zn2+, Cu2+, Ca2+, Mg2+, Mn2+, and Fe2+ inhibited the enzyme by 20% to 80%. An investigation was also done on the influence of organic solvents like methanol, ethanol, acetone, and isopropyl alcohol on enzyme stability. The results showed that the enzyme was stable in all the solvents, exhibiting 43% to 70% relative activity. The enzyme kinetic studies indicated Km and Vmax values of 4.54 mg ml-1 and 76.92 UmL-1, respectively. The enzyme also demonstrated antifungal activity against Aspergillus niger NCIM 1207 using the agar well diffusion method.
Keywords: Bacillus cereus, Enzyme purification, Chitinase characterization, Enzyme kinetics, Antifungal activity
Introduction
Chitin, a supportive, protective, insoluble polysaccharide (β-1,4-N-acetyl-D-glucosamine) profusely present as a structural constituent in mollusk shells, exoskeleton of arthropods, and fungal cell walls. As a biopolymer, it comes in second globally after cellulose producing more than 1010 metric tons per year primarily derived from marine waters alone (Hao et al., 2012; Loni et al., 2014). It is essential to recycle chitin into nature, and this can be achieved by a group of enzymes, namely, lysosomes and chitinases (Korany et al., 2019). Chitinases (EC 3.2.1.14) are the enzymes that hydrolyze homopolymer links of β-1,4-N-acetyl-D-glucosamine into monomer N-acetyl glucosamine (GlcNAc) (Loni et al., 2014). Nutrient uptake, structural changes, defense, and attack are some of the biological functions in which these broadly distributed enzymes take a vital part (Loni et al., 2014). Chitinases are produced by bacteria, fungi, actinomycetes, higher plants, lower animals, and birds (Ghorbel-Bellaaj et al., 2012; Lestari et al., 2017; Cheba et al., 2018). Carbon and nitrogen are sourced from chitin by microorganisms (Lestari et al., 2017; Aktuganov et al., 2018). Microorganisms produce chitinase because they use chitin as an energy source, while fungi and insects produce chitinase because they are involved in morphogenesis (Korany et al., 2019). Among prokaryotes, actinomycetes are the major group producing potent chitinases (Ghorbel-Bellaaj et al., 2012). The main genus of this group is Streptomyces; some chitinases are produced by several species such as S. aureofaciens, and S. olivaceoviridis for resourceful degradation of chitin. Other actinomycetes that produce chitinase include Actinoplanes, Micromonospora, and Nocardia (Aktuganov et al., 2018; Cheba et al., 2018; Ray et al., 2019; Akeed et al., 2020).
The classification of the Chitinase family is done as per the action mechanism i.e., individual hydrolysis mode of the enzyme and homology of their amino acid sequences. Generally, chitinases are randomly splitting hydrolases that recognize the O-glycosyl bond between Chito-saccharide residues for catalysis. There are two classes of chitinase enzymes, namely, endo-chitinases and exo-chitinases (Ray et al., 2019). The hydrolysis of internal O-glycosyl bonds to cleaving the polymers of chitin into oligosaccharides is done by the endo-chitinases, whereas the cleaving of only terminal GlcNAc residues is performed by the exo-chitinases from the chitin polymers’ reducing end. The decomposition of chitin polymers into soluble N-acetylglucosamine units is possible due to the dual action of the exo- and endo-chitinases (Ray et al., 2019). There are many reports on the production of chitinases with diverse molecular masses by various bacterial genera, including Bacillus (Lestari et al., 2017). Several species of Bacillus such as B. pumilus, B. circulans, B. subtilis, Virgibacillus marismortui, B. licheniformis, B. cereus, Bacillus subtilis, B. thuringiensis, etc. are known to produce chitinase enzyme (Loni et al., 2014; Akeed et al., 2020).
Chitinases find various industrial and environmental applications. They are vital in the geochemical recycling of carbon and nitrogen and give them potential in pest control, bioremediation, and various other biotechnological applications (Loni et al., 2014). The enzyme was used to produce protoplasm when applied to fungal and yeast cells by degrading their cell walls indicating their potential application in the biological control of plant pathogens (Loni et al., 2014; Ray et al., 2019). Chitinases can also be used in the management of marine waste (Hao et al., 2012) and in the production of bioactive derivatives like chitooligosaccharides and functional foods (Le & Yang, 2019).
Chitosan and other derivatives of the enzymes are also increasingly being used in several industries, and enzymatic methods of obtaining them are becoming increasingly important. Traditionally, the chitin was degraded by various chemical and physical methods, which resulted in their low efficiency, high cost, and lack of specificity even though they have been industrialized (Meriem & Mahmoud, 2017). These limitations can be overcome by an eco-friendly enzymatic degradation. As the degradation kinetics of chitinases is relatively slower, the industrial applications of chitinases are limited (Meriem & Mahmoud, 2017; Rohmani et al., 2022). Hence, more efforts and studies are needed to explore newer enzyme sources, adopt better production strategies, and the invention of newer chitinase forms with a wider range of functionality.
In this study Bacillus cereus, BSH-04 was used to produce chitinase. Ammonium sulfate precipitation and dialysis were carried out for the partial purification of the enzyme, whereas the molecular weight of the enzyme was determined by SDS-PAGE and silver staining method. The effect of various parameters such as pH, temperature, metal ions, various solvents, and various concentrations of a substrate on chitinase activity was investigated. The antifungal activity of the enzyme was performed using the agar well diffusion method against Aspergillus niger NCIM 1207.
Materials and Methods
Isolation and Screening of Chitinolytic Bacteria
The bacterial strain of Bacillus cereus BSH-04 was isolated from a marine soil source. The strain was isolated using basal media containing KH2PO4 (0.1%), Na2HPO4 (0.2%), CaCl2. 7H2O (0.05%), MgSO4.7H2O (0.05%), yeast extract (0.25%), NH4Cl 0.1%, NaCl 0.05%, colloidal chitin (1%), incubated for 96 hrs at 30ºC (Shaikh et al., 2018). The chitinolytic activity of the strain was checked by screening the isolate on an agar plate containing 0.5% colloidal chitin as substrate at room temperature with an incubation time of 48 h to 72 hrs.
Molecular Identification of the Bacterial Isolate by 16s Rrna Gene Sequencing
The whole genomic DNA was extracted from the pure culture using the ionic detergent cetyl trimethyl ammonium bromide (CTAB) method. Amplification of 16S fragment (~1300-1500bp) was performed by High-fidelity polymerase using a thermocycler. The amplified fragment was then purified and subjected to nucleotide sequencing. The fragment was sequenced by primers, 63F (5'CAGGCCTAACACATGCAAGTC3') and 1387R (5'GGCGGATGTGTACAAGGC3'). The sequencing was carried-out using big dye terminator version 3.1 analyzed by ABI 3130 Genetic Analyser. Using the MEGA 6.0 tool, a phylogenetic tree was constructed by the neighbor-joining method.
Colloidal Chitin Preparation
Colloidal chitin used as a substrate for chitinase assay was prepared by dissolving 10 g of chitin flakes in 60 mL of concentrated HCl under constant stirring for 1 to 2 hrs at room temperature. The solution was then poured into 400 mL of deionized water with continuous stirring to form a precipitate. Further centrifugation was employed to collect it and attained the neutral pH after several washes by water thus, obtained colloidal chitin was stored at 4ºC until further use (Jha et al., 2016; Bouacem et al., 2018; Vasetska et al., 2022).
Assay of Chitinase Activity
Protein was estimated according to the Lowry method using bovine serum albumin as a standard (Shehata et al., 2018). A spectrophotometer was utilized in the measurement of the chitinolytic activity through the determination of the free groups formed by chitin hydrolysis. The reaction mixture consisted of sodium phosphate buffer (0.2M, pH 7.0) that was used to suspend 1 ml of colloidal chitin (0.5%). The whole content was incubated for 30 min at 37oC, and the reaction was terminated by the addition of 1 mL DNS. Further, the content was heated in boiling water for 5 min and the absorbance of the supernatant was measured at 540 nm. One unit of chitinase activity was defined as the production of 1 µmol (NAG) per minute using a standard curve (N-acetyl glucosamine) (Bouacem et al., 2018; Batarseh et al., 2022).
Profiling Study for Growth and Chitinase Production
Bacterial growth and chitinase production were studied by inoculating the culture at log-phase into a production medium containing colloidal chitin (pH 7). Further, it was incubated at 30ºC under constant shaking at 120 rpm. Samples were collected every 24 h for five days. The absorbance at 600 nm from a UV-Visible spectrophotometer was used to measure the culture growth. The growth curve was plotted by considering the incubation time on the X-axis and absorbance on the Y-axis. Each sample was analyzed for enzyme activity by the method explained above (Bouacem et al., 2018). Top of Form
Partial Purification of Chitinase
The enzyme present in the cell-free supernatant was partially purified by ammonium sulfate precipitation at different saturation concentrations like 70%, 75%, 80%, and 85% at 4°C under constant stirring. The precipitated solution was subjected to centrifugation at 8000 rpm for 20 min at 4°C. The precipitate was again dissolved in 4 ml of sodium acetate buffer (pH 7.0). Consecutively, the crude protein was further subjected to dialysis against the same buffer for 6h at 4°C and the buffer was changed every 3 hrs. The protein content and chitinase activity in each fraction were determined (Kavitha & Vijayalakshmi, 2011).
Molecular Weight Determination by SDS-PAGE
The molecular weight of the partially purified chitinase enzyme was determined by the preference fraction with the highest chitinase activity. Sodium dodecyl sulfate gel electrophoresis (SDS-PAGE) was used to evaluate the molecular weight (Ray et al., 2019; Remizova et al., 2022), containing 5% and 10% of acrylamide for stacking gel and separating gel, respectively. For protein identification, a sensitive staining method i.e., the silver staining method, was used to detect the molecular weight of partially purified chitinase enzyme (Senol et al., 2014; Kumar et al., 2022).
Characterization of Partially Purified Chitinase
Effect of pH on Enzyme Activity
The effect of pH on chitinase was identified by incubating the reaction mixture with various pH values from 4 to 10, such as 0.1 M acetate buffer (pH 4–6), 0.1 M phosphate buffer (pH 5–7), and 0.1 M Tris buffer (pH 6.5-10) for 30 min at 37°C. The pH stability for chitinase was determined by incubating the enzyme without substrate in the presence of various pH (4 to 10) for 1h at 4°C, and the relative activity was measured using colloidal chitin as a substrate (Liu et al., 2010; Cheba et al., 2016).
Effect of Temperature on Enzyme Activity
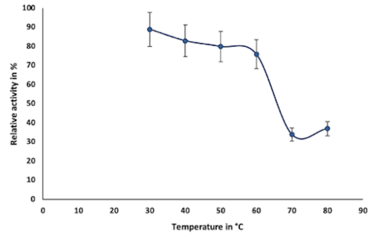
To study the effect of temperature on enzyme activity, we varied the temperature from 10-70oC and performed enzyme assay by standard method. The thermal stability of the enzyme was determined by pre-incubating the enzyme at various temperature ranges (30⁰C to 80°C) without substrate in the reaction buffer at pH 6.0 for 1 h, later relative activity was calculated by using colloidal chitin as substrate under standard conditions (Liu et al., 2010; Cheba et al., 2016; Maghami et al., 2022).
Effect of Various Metal Ions on Enzyme Activity
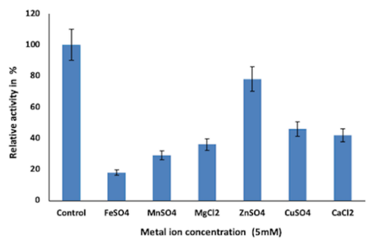
To investigate the effect of different metal ions on enzyme activity metal ions (Zn2+, Cu2+, Ca2+, Mg2+, Mn2+, and Fe2+) were added to the reaction mixture. The final concentration attained was 5 mM and further Chitinolytic activity was tabulated using standard assay protocols.
Effect of Organic Solvents on Enzyme Activity
The study of the influence of various organic solvents on purified chitinase was performed by incubating the enzyme with different solvents, including methanol, ethanol, and acetone, isopropyl alcohol, with a final concentration of 10% and 20%, respectively.
Effect of Substrate Concentration on Enzyme Activity
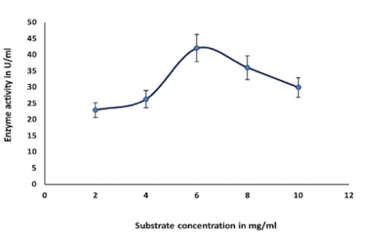
Colloidal chitin was utilized in the study of substrate affinity. The affinity between substrate and enzyme was determined by incubating the purified chitinase with various colloidal chitin (2 mg mL-1 to 10 mg mL-1). Lineweaver- Burk (LB) curve was used to calculate Km and Vmax (Doan et al., 2021; AlShehri et al., 2022).
Antifungal Activity of the Partially Purified Enzyme
The antifungal activity of the partially purified chitinase was tested against Aspergillus niger NCIM 1207 using the agar well diffusion method (Delfini et al., 2021; AlHumaidi et al., 2022; Burlaka, 2022). The fungus was seeded (100 μL) onto the potato dextrose agar medium (PDA). Then, 6 mm diameter holes were pressed aseptically, loaded with 100 μL of the partially purified chitinase, and then incubated for four days at 30°C. The antifungal activity was evaluated by measuring (in mm) the inhibition zone of the fungal growth around the hole.
Results and Discussion
Screening of Chitinase Producing Organism
In the current study, a bacterial strain was identified as a chitinase producer based on the formation of clear zones on an agar plate containing 0.5% colloidal chitin and culture at 30°C for 72 hrs. The zone of clearance around the bacterial culture showed extracellular chitinase activity after 72 hrs of incubation in the optimized medium.
Molecular Identification by 16s rRNA Gene Sequencing
Identification of strain was achieved by comparing the 16S rRNA gene sequences of the strain with 16S rRNA gene sequences of the NCBI GenBank database with the help of BLAST; the isolate was found to be Bacillus cereus strain BSH-04 (GenBank Accession No. MT672784.1) by analyzing the phylogenetic tree which gave the information related to the highest identity of strain and its evolutionary relationships which was shown in Figure 1.
|
|
|
Figure 1. Phylogenetic analysis of Bacillus cereus BSH-04 |
Profiling Study for Growth and Enzyme Production
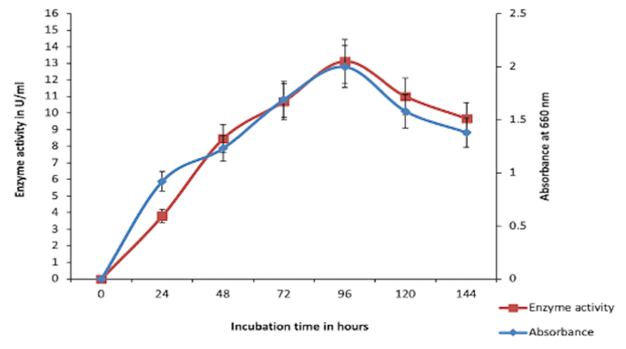
The bacterial growth and chitinase production were increased from 24 to 96 hrs of incubation and reached a maximum at 96 h. As shown in Figure 2, the highest enzyme activity was observed after 96 hrs of incubation later, bacterial growth reached a stationary phase, and enzyme activity decreased.
|
|
|
Figure 2. Profiling study for growth and chitinase production by Bacillus cereus BSH-04 |
Purification of chitinase by ammonium sulfate precipitation and dialysis
The precipitation of chitinase at diverse ammonium sulphate saturation was performed. The results showed that at 70%, 75%, 80% and 85% saturation enzyme activities reported were 108.67 U/ml, 107.45 U/ml, 143.68 U/ml, and 133.33 U/ml respectively. The highest protein and maximum chitinase activity were observed to be 1.50 mg ml-1 and 143.68 UmL-1, respectively at 80% ammonium sulfate saturation; further dialysis resulted in an increase in the purification fold up to 1.75 along with maximum chitinase activity (288.88 UmL-1) was shown in Table 1.
Table 1. Purification table for chitinase enzyme.
|
Sample name |
Total activity (U) |
Total protein (mg) |
Specific activity (U/mg) |
Purification fold |
|
Crude |
43763.9 |
305.95 |
143.04 |
1 |
|
Dialyzed |
1068.56 |
4.255 |
251.13 |
1.75 |
Molecular Weight Determination by SDS PAGE and Silver Staining
The sodium dodecyl sulfate gel electrophoresis (SDS-PAGE) was employed to assess the enzyme molecular weight which was observed to be 60 kDa. The marker used was 10 kDa to 100 kDa; due to lower loading, sample silver staining was carried out, which is shown in Figure 3.
|
|
|
Figure 3. Figure showing molecular weight of partially purified chitinase using SDS-PAGE method |
Characterization of Partially Purified Chitinase
Effect of pH on Enzyme Stability and Activity
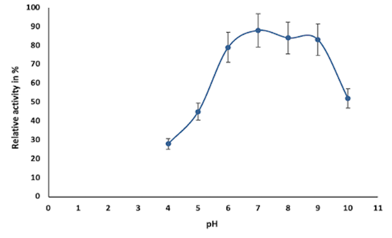
Figure 4a and Figure 4b provide the effect of pH on enzyme stability and activity, respectively. Chitinase activity was less at pH 4.0 and gradually increased from pH 5 to 7 and attained optimum at pH 7.0; further activity decreased from pH 8 to pH 10, which was shown in Figure 4a. The stability of chitinase was found in the range of pH 6.0 to 9.0 (Figure 4b).
Effect of Temperature on Enzyme Stability and Activity
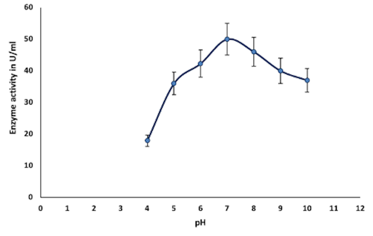
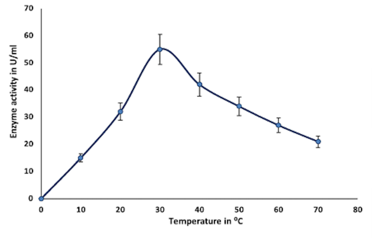
Enzymes are proteins in nature, inactivates above 50°C, denatures at 70°C, and are unable to provide sufficient activation energy at very lower temperatures (El-Sayed et al., 2019). In this study, the optimum temperature for the purified enzyme was observed at 30°C, which was earlier reported as 37°C by (El-Sayed et al., 2019); after attaining optimum temperature, chitinase activity gradually decreases from 40°C to 70°C (Figure 4c). The stability of chitinase was found in the range of 30°C to 60°C (Figure 4d).
Effect of Various Metal Ions on Enzyme Activity
Figure 4e shows the influence on enzyme activity from various metal ions. Inhibition of enzyme at 5mM concentration of all metal ions (Zn2+, Mg2+, Mn2+, Cu2+, Ca2+, and Fe2+) was observed. Zn2+ showed lesser inhibition, whereas Fe2+ showed the highest enzyme inhibition; overall enzyme inhibition was from 20% to 80%.
Effect of Organic Solvents on Enzyme Activity
The values for relative enzyme activity with 10% each of ethanol, methanol, acetone and isopropyl alcohol were found to be 47%, 43%, 70%, and 53% respectively. Hence, it is observed that the enzyme is more stable in acetone and less stable in methanol.
Effect of Substrate Concentration
Enzyme activity was found to be optimum at 6 mg ml-1 of colloidal chitin and decreased in the range of 2, 4,8,10 mg ml-1 (colloidal chitin), which is shown in Figure 4f.
|
|
|
a) |
|
|
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
|
|
e) |
|
|
|
f) |
|
Figure 4. a, b, c, d) show the effect and stability of chitinase enzyme at different pH and temperature values respectively. E, f) show the effect of different metal ions (5mM) and substrate concentration respectively |
Determination of Km and Vmax
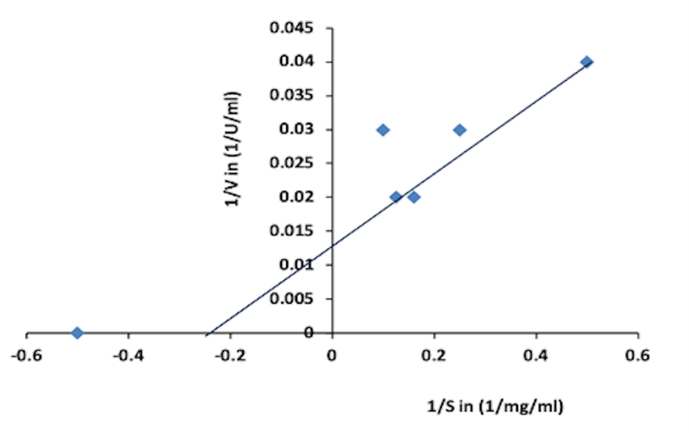
Vmax and Km were respectively found to be 76.92 Umg-1 and 4.54 mg ml-1, from the Lineweaver-Burk plot shown in Figure 5 here the small Km indicates a high affinity of the enzyme towards the colloidal chitin, Km and Vmax kinetic constants of purified chitinase from Paenibacillu spasadenensis CS0611 were reported as 4.41 mg ml-1 and 1.08 mg min-1 respectively by (Guo et al., 2017; Dhanasekar et al., 2022); similarly, Km and Vmax of Bacillus pumilus JUBCH08 were reported as 0.13 mg ml-1 and 38.23 U ml-1 respectively by (Bhattacharya et al., 2016).
|
|
|
Figure 5. Lineweaver Burk plot (The Km and Vmax obtained as 4.54 mg/ml and 76.92 U/mg respectively) |
Antifungal Activity
Antifungal activity against Aspergillus niger NCIM 1207 was tested by the agar well diffusion method. The zone of inhibition (ZoI) was observed when the partially purified chitinase was added to the PDA agar seeded with fungal spores incubated for 96 h at 30°C. No zone of inhibition was observed in the negative control. In agriculture, the chitinases are used for the biological control of pests and plant fungal pathogens such as Trichoderma reesei, Colletotrichum gleosporoides, Phoma medicaginis, Rhizoctonia solani, etc (Oyeleye & Normi, 2018). Similar fungal inhibition was reported against sixteen fungal strains by chitinase enzyme present in the cell-free extract in a previous report (Delfini et al., 2021). The researchers had reported inhibition of hyphae development and germination of spores by cell-free enzyme preparation.
Last few decades, the production of microbial chitinase has attracted attention. Many organisms have been reported to produce chitinase enzyme; the bacteria are the major source of chitinase-producing microorganisms (Bhattacharya et al., 2007). In the present study, the bacterial strain Bacillus cereus BSH-04 was isolated from a marine soil source and further used for enzyme production. The profiling study of the bacterial growth and chitinase production from Bacillus cereus BSH-04 was studied from 24 hrs to 144 hrs of incubation. From Figure 2, a steady-steep growth of bacteria was observed around 0 h to 24 hrs, whereas the production of chitinase was less rapid in the initial growth period of 24 hrs. From the onset of 24 h to 96 hrs, a steady increase in slope was observed (bacterial growth), whereas the enzyme production was also increased. The highest growth and chitinase production was also observed at 96 hrs with maximum chitinase activity of 13.13 UmL-1. Thereafter, declination of the bacterial growth along with chitinase production was observed till 144 hrs. It can be inferred that chitinase is a growth-dependent enzyme, and the production of the enzyme depreciates as the cell undergoes into stationary phase and death phase. In this phase, the chitinolytic activity gradually decreases, which may be due to the accumulation of by-products resulting in a negative feedback mechanism (Khan et al., 2015).
Partial purification of chitinase enzyme from the supernatant at 80% ammonium sulfate saturation exhibited the highest protein recovery with the highest activity (143.68 UmL-1). Similar results have been reported by earlier researchers (Hammami et al., 2013; Shivakumar et al., 2014). Further purification, i.e., dialysis, has increased the purification fold to 1.75 with a specific activity of 251.13 Uml-1. The sodium dodecyl sulfate gel electrophoresis (SDS-PAGE) and silver staining methods were employed to assess the enzyme molecular weight observed to be 60 kDa. Various bacterial genera (Bacillus, Streptomyces, Brevibacillus, Serratia, Chromobacterium, etc) were reported to produce chitinase with molecular weights around 60-65 kDa (Poria et al., 2021). Similar reports have been reported by (Bhattacharya et al., 2016; Singh et al., 2021). In a similar study, the molecular weight of chitinase from Paenibacillus pasadenensis CS0611was found to be 69 kDa (Guo et al., 2017). Since the chitinase enzyme is a protein, high temperatures can inactivate or denature the enzyme. This may be due to hydrolysis of the peptide chain, amino acid aggregation, incorrect confirmation, or disruption of amino acids (Shaikh et al., 2018).
Generally, the pH optima for most chitinases were in the acidic range. Hence, changes in the pH value of the medium affect enzyme stability and activity as well. This could be due to the characteristic poly-extremophiles of bacteria present in particular ecosystems (Aktuganov et al., 2018). Metal ions function as links to both the substrate and the enzyme and the substrate, binding to them both thus stimulating enzyme activity by keeping the substrate in the active site of the enzyme (Shivakumar et al., 2014). The effect of metal ions on enzyme activity can also be attributed to the changes in electrostatic bonds that can alter the tertiary structure of the enzyme. This may also be because the sulfhydryl group may be involved in the active site of the enzyme (Shivakumar et al., 2014). Figure 4e indicates that the residual activity of chitinase from Bacillus cereus BSH-04 was inhibited from 20% to 80% in the presence of metal ions like Zn2+, Cu2+, Ca2+, Mg2+, Mn2+ and Fe2+ at 5 mM concentration. Fe2+ inhibited the chitinase activity to a greater extent, i.e., by almost 80%, and least inhibited by Zn2+, i.e., 20%. Similar results were reported by (Hao et al., 2012; Shivakumar et al., 2014). In contrast, Fe and Mg in (Ghorbel-Bellaaj et al., 2012) and Mg, Ca, and Cu in (Bouacem et al., 2018) have increased the relative chitinase activity. In (Aktuganov et al., 2018), Ca and Mg neither showed any significant inhibitory or activating effects on the enzyme. Enzyme inhibition by Zn may be associated with residues of acids like aspartic and glutamic acids left in chitinase. studies have indicated that these amino acids can bind to specific divalent cations in the active site of chitinase, thereby inhibiting chitinase (Shivakumar et al., 2014).
Our studies showed that the chitinase exhibited a moderate tolerance to some organic solvents such as methanol, ethanol, acetone, and isopropanol. The enzyme activity was inhibited at around 50% in the presence of 10% ethanol, methanol, and isopropyl alcohol. Therefore it can be concluded that the enzyme stability across the given solvents indicates the suitability of the enzyme in industrial applications.
Lineweaver-Burk plots were used in the determination of the kinetic values of Km and Vmax, whose values were found to be 4.54 mg mL-1 and 76.92 Umg-1, respectively (Figure 5). The kinetic parameters Km and Vmax define substrate specificity. Lower Km and higher Vmax values verify the chitinase enzyme specificity to colloidal chitin (Shehata et al., 2018). Such investigations indicate the suitability of the chitinase enzyme in many biochemical transformations.
Members of the genus Bacillus are also known to produce a wide variety of antibacterial compounds, the most biologically active molecules being non-ribosomal and synthetic lipopeptides. B. amyloliquefaciens PGP BacCA1 cells showed antifungal effects against some phytopathogenic fungi when applied to bean seeds (Delfini et al., 2021). An important observation from this study is the sprouting inhibition shown for A. niger NCIM 1207 by partially purified chitinase, as spore germination is the first step in the activation of the asexual life cycle and the spread of multiple phytopathogenic air fungal diseases. Therefore, if the plant pathogens are inhibited at this stage, this disease can be effectively controlled (Delfini et al., 2021).
Conclusion
Production of chitinase is considered to be an important matter as the recycling of insoluble chitin into its simpler forms. The microbial production of chitinase is considered to be more efficient; therefore, the enzyme production from Bacillus cereus BSH-04 was undertaken for the study. The partial purification of 60kDa chitinase resulted in an increase in the purification fold by 1.75. The enzyme demonstrated temperature and pH stability that may be useful in various industrial applications of the enzyme at different temperatures and pH values. The study also depicted various kinetic parameters which are useful to understand the stability of chitinase enzymes under different physicochemical conditions. The antifungal activity as observed against Aspergillus niger indicates the use of chitinase both as an antifungal and biocontrol agent.
Acknowledgments: None
Conflict of interest: None
Financial support: None
Ethics statement: None
Akeed, Y., Atrash, F., & Naffaa, W. (2020). Partial purification and characterization of chitinase produced by Bacillus licheniformis B307. Heliyon, 6(5), e03858.
Aktuganov, G. E., Galimzianova, N. F., Gilvanova, E. A., Kuzmina, L. Y., Boyko, T. F., Safina, V. R., & Melentiev, A. I. (2018). Characterization of Chitinase Produced by the Alkaliphilic Bacillus mannanilyticus IB-OR17 B1 Strain. Applied Biochemistry and Microbiology, 54(5), 505-511.
AlHumaidi, M. A., AlJahdali, S. L., AlGab, M. K. M., AlRizqi, A. A., AlShammeri, A. N., & AlQarni, A. A. (2022). A survey-based study on dentists' knowledge and practices regarding lasers in Riyadh City. Annals Journal of Dental and Medical Assisting, 2(1), 8-13. doi:10.51847/E6TbI0HfXU
AlShehri, O. M., Jali, N. M., Almutairi, Y. M., Aljrais, M. M., Alsirhani, A. M., & AlQudairi, A. S. (2022). Common causes of cusp fracture in adults; a systematic review. Annals of Dental Specialty, 10(2), 35-38. doi:10.51847/jBTdUqt7Cx
Batarseh, N., Khalil, R., & Al-Domi, H. A. (2022). Hypothalamic neuroinflammation induced by obesity and the effect of Liraglutide. Journal of Advanced Pharmacy Education and Research, 12(1), 46-55. doi:10.51847/N1hvOB1anS
Bhattacharya, D., Nagpure, A., & Gupta, R. K. (2007). Bacterial chitinases: properties and potential. Critical Reviews in Biotechnology, 27(1), 21-28.
Bhattacharya, S., Das, A., Samadder, S., & Rajan, S. S. (2016). Biosynthesis and characterization of a thermostable, alkali-tolerant chitinase from Bacillus pumilus JUBCH08 displaying antagonism against phytopathogenic Fusarium oxysporum. 3 Biotech, 6(1), 1-8.
Bouacem, K., Laribi-Habchi, H., Mechri, S., Hacene, H., Jaouadi, B., & Bouanane-Darenfed, A. (2018). Biochemical characterization of a novel thermostable chitinase from Hydrogenophilus hirschii strain KB-DZ44. International Journal of Biological Macromolecules, 106, 338-350.
Burlaka, I. (2022). Approach to correction of apoptotic disorders in children with early diabetic nephropathy. Journal of Advanced Pharmacy Education and Research, 12(2), 104-109. doi:10.51847/G6i3231Jnm
Cheba, B. A., Zaghloul, T. I., EL-Mahdy, A. R., & EL-Massry, M. H. (2016). Effect of pH and temperature on Bacillus sp. R2 chitinase activity and stability. Procedia Technology, 22, 471-477.
Cheba, B. A., Zaghloul, T. I., El-Mahdy, A. R., & El-Massry, M. H. (2018). Effect of nitrogen sources and fermentation conditions on Bacillus sp. R2 chitinase production. Procedia Manufacturing, 22, 280-287.
Delfini, C. D., Villegas, L. B., Martínez, M. A., & Baigorí, M. D. (2021). Extracellular Antifungal Activity of Chitinase-Producing Bacteria Isolated From Guano of Insectivorous Bats. Current Microbiology, 78(7), 2787-2798.
Dhanasekar, P., Rajayyan, J. S., Veerabadiran, Y., Kumar, K. S., Kumar, K. S., & Chinnadurai, N. (2022). Evaluation of alum and purification process of water by coagulation method. Bulletin of Pioneering Researches of Medical and Clinical Science, 1(2), 1-6. doi:10.51847/R8GyfOmMDh
Doan, C. T., Tran, T. N., & Wang, S. L. (2021). Production of Thermophilic Chitinase by Paenibacillus sp. TKU052 by Bioprocessing of Chitinous Fishery Wastes and Its Application in N-acetyl-D-glucosamine Production. Polymers, 13(18), 3048.
El-Sayed, M., Nassar, O., Nasr, H., & Kobisi, A. E. N. (2019). Efficacy of thermophilic soil-isolated Paenibacillus sp. NBR10 as a chitinolytic and biocontrol bacterium-in vitro study. Egyptian Journal of Botany, 59(1), 195-208.
Ghorbel-Bellaaj, O., Manni, L., Jellouli, K., Hmidet, N., & Nasri, M. (2012). Optimization of protease and chitinase production by Bacillus cereus SV1 on shrimp shell waste using statistical experimental design. Biochemical and molecular characterization of the chitinase. Annals of Microbiology, 62(3), 1255-1268.
Guo, X., Xu, P., Zong, M., & Lou, W. (2017). Purification and characterization of alkaline chitinase from Paenibacillus pasadenensis CS0611. Chinese Journal of Catalysis, 38(4), 665-672.
Hammami, I., Siala, R., Jridi, M., Ktari, N., Nasri, M., & Triki, M. A. (2013). Partial purification and characterization of chi IO 8, a novel antifungal chitinase produced by B acillus cereus IO 8. Journal of Applied Microbiology, 115(2), 358-366.
Hao, Z., Cai, Y., Liao, X., Zhang, X., Fang, Z., & Zhang, D. (2012). Optimization of nutrition factors on chitinase production from a newly isolated Chitiolyticbacter meiyuanensis SYBC-H1. Brazilian Journal of Microbiology, 43, 177-186.
Hehata, A. N., Abd El Aty, A. A., Darwish, D. A., Wahab, W. A. A., & Mostafa, F. A. (2018). Purification, physicochemical and thermodynamic studies of antifungal chitinase with production of bioactive chitosan-oligosaccharide from newly isolated Aspergillus griseoaurantiacus KX010988. International Journal of Biological Macromolecules, 107, 990-999.
Jha, S., Modi, H. A., & Jha, C. K. (2016). Characterization of extracellular chitinase produced from Streptomyces rubiginosus isolated from rhizosphere of Gossypium sp. Cogent Food & Agriculture, 2(1), 1198225.
Kavitha, A., & Vijayalakshmi, M. (2011). Partial purification and antifungal profile of chitinase produced by Streptomyces tendae TK-VL_333. Annals of Microbiology, 61(3), 597-603.
Khan, F. I., Bisetty, K., Singh, S., Permaul, K., & Hassan, M. (2015). Chitinase from Thermomyces lanuginosus SSBP and its biotechnological applications. Extremophiles, 19(6), 1055-1066.
Korany, S. M., Mansour, A. N., El-Hendawy, H. H., Kobisi, A. N. A., & Aly, H. H. (2019). Entomopathogenic efficacy of the chitinolytic bacteria: Aeromonas hydrophila isolated from Siwa Oasis, Egypt. Egyptian Journal of Biological Pest Control, 29(1), 1-10.
Kumar, D., Gurunathan, D., Jabin, Z., & Talal, S. (2022). Comparative efficacy of aromatherapy and conscious sedation in pediatric dental anxiety management. Annals Journal of Dental and Medical Assisting, 2(1), 14-21. doi:10.51847/TVRFhVaiIQ
Le, B., & Yang, S. H. (2019). Microbial chitinases: properties, current state and biotechnological applications. World Journal of Microbiology and Biotechnology, 35(9), 1-12.
Lestari, P., Prihatiningsih, N., & Djatmiko, H. A. (2017, February). Partial biochemical characterization of crude extract extracellular chitinase enzyme from Bacillus subtilis B 298. In IOP Conference Series: Materials Science and Engineering (Vol. 172, No. 1, p. 012041). IOP Publishing..
Liu, D., Cai, J., Xie, C. C., Liu, C., & Chen, Y. H. (2010). Purification and partial characterization of a 36-kDa chitinase from Bacillus thuringiensis subsp. colmeri, and its biocontrol potential. Enzyme and Microbial Technology, 46(3-4), 252-256.
Loni, P. P., Patil, J. U., Phugare, S. S., & Bajekal, S. S. (2014). Purification and characterization of alkaline chitinase from Paenibacillus pasadenensis NCIM 5434. Journal of Basic Microbiology, 54(10), 1080-1089.
Maghami, H., Parhizkar, H., Riasaty, A., Banani, A., Poustfroosh, M., & Hasehmpour-Sadeghian, M. (2022). Identifying the parenting style of students’ parents of an school at Shiraz University of medical sciences. Journal of Advanced Pharmacy Education and Research, 12(1), 91-96. doi:10.51847/tLcoO2yvKI
Meriem, G., & Mahmoud, K. (2017). Optimization of chitinase production by a new Streptomyces griseorubens C9 isolate using response surface methodology. Annals of Microbiology, 67(2), 175-183.
Oyeleye, A., & Normi, Y. M. (2018). Chitinase: diversity, limitations, and trends in engineering for suitable applications. Bioscience Reports, 38(4)., 1-21.
Poria, V., Rana, A., Kumari, A., Grewal, J., Pranaw, K., & Singh, S. (2021). Current perspectives on chitinolytic enzymes and their agro-industrial applications. Biology, 10(12), 1319.
Ray, L., Panda, A. N., Mishra, S. R., Pattanaik, A. K., Adhya, T. K., Suar, M., & Raina, V. (2019). Purification and characterization of an extracellular thermo-alkali stable, metal tolerant chitinase from Streptomyces chilikensis RC1830 isolated from a brackish water lake sediment. Biotechnology Reports, 21, e00311.
Remizova, A. A., Bitarov, P. A., Epkhiev, A. A., & Remizov, N. O. (2022). Reparative-regenerative features of bone tissue in experimental animals treated with titanium implants. Journal of Advanced Pharmacy Education and Research, 12(2), 110-116. doi:10.51847/Sprxb1DKyv
Rohmani, S., Desi, B. A., & Wardhani, W. D. (2022). Potassium-Azeloyl-Diglycinate BB-Cream formulation with Triethanolamine variation, and its effects on In-Vitro SPF stability and values. Journal of Advanced Pharmacy Education and Research, 12(1), 1-8. doi:10.51847/MUHp4ptold
Senol, M., Nadaroglu, H., Dikbas, N., & Kotan, R. (2014). Purification of Chitinase enzymes from Bacillus subtilis bacteria TV-125, investigation of kinetic properties and antifungal activity against Fusarium culmorum. Annals of Clinical Microbiology and Antimicrobials, 13(1), 1-7.
Shaikh, S. S., Wani, S. J., & Sayyed, R. Z. (2018). Thakur R, Gulati A. Production, purification and kinetics of chitinase of Stenotrophomonas maltophilia isolated from rhizospheric soil. Indian Journal of Experimental Biology, 56, 274-278.
Shivakumar, S., Karmali, A. N., & Ruhimbana, C. (2014). Partial purification, characterization, and kinetic studies of a low-molecular-weight, alkali-tolerant chitinase enzyme from Bacillus subtilis JN032305, a potential biocontrol strain. Preparative Biochemistry and Biotechnology, 44(6), 617-632.
Singh, R., Upadhyay, S. K., Singh, M., Sharma, I., Sharma, P., Kamboj, P., Saini, A., Voraha, R., Sharma, A. K., Upadhyay, T. K., et al. (2021). Chitin, chitinases and chitin derivatives in biopharmaceutical, agricultural and environmental perspective. Biointerface Research in Applied Chemistry, 11(3), 9985-10005.
Vasetska, O., Zhminko, P., Prodanchuk, M., Galkin, A., & Tsygankova, V. (2022). Perspective for using 2,6-dimethylpyridine-N-oxide to reduce the toxic effect of xenobiotics in mammals. Journal of Advanced Pharmacy Education and Research, 12(1), 21-29. doi:10.51847/TXCxI0PsO1