
Grass and Legume Plant Productivity in Spring and Summer Planting
Nikolay Buyankin, Andrey Krasnoperov*
Abstract
Planting a mixture of grass and legume seeds with different linear growth rates allows one to set multilayer crops where the bottom layer is taken by lupine and millet, and the top one – by beans, vetch, and peas. This distribution allows the plants to maximize their sunlight intake and prevents soil moisture evaporation. Analysis of the rhizospheric indices has made it possible to determine the way soil rhizospheric processes develop in legume crops after the spring and summer plantings. In the spring mixtures, they shifted рНKCl to be more acetous increasing the amount of available chemical elements in the rhizosphere, unlike the summer mixture making рНKClmore alkali and increasing the amounts of nitrogen, phosphorous and potassium in the rhizosphere. Number of ammonifying bacteria after mixed summer planting increased 1.7-2.1 times if compared to that after spring planting, which was due to more favorable conditions when more of the easily hydrolyzed rhizospheric compounds were produced.
Key words: grass and legume crops, microbiological soil activity, high-protein feed, soil fertility, dry grass yield
Introduction
In most of the corn-breeding regions of the Russian Federation, the second part of the vegetation period (July-August) is more favorable in terms of temperature and precipitation, so summer crops are normally able to form up to 60-80 t of their herbage by October. Being plowed under, this herbage facilitates increasing the mobile part of fresh organic matter and reduces soil degradation (Benc, 1999; Zvyagintsev et al. 2005).
Since 1993 in the sod-podzolic, mildly argillaceous, and moderately cultivated soil of the Kaliningrad Region, an experiment has been going on to compare the summer and spring long-day annuals (Krasnoperov, 2019). Recently, in the field experiments, one has been studying different seed mixtures including lupin, field beans, spring and autumn vetch, and millet. The obtained multiyear data on herbage yield have confirmed the effectiveness of summer planting in the environmental conditions of the Kaliningrad Region.
Literature review
Grass and legume crops are the main source of high-quality food products and forage. Legume crops also facilitate reducing CO2emissions, since they produce 5-7 times less CO2 per unit area than other plants (Clune et al., 2017, Stagnariet al., 2017), which may have a rather positive effect in terms of global warming (Guardia et al., 2016).
It is a well-known fact that grass and legume crops affect soil microflora that may include fungi and different kinds of bacteria such as psychrotrophic, mesophilic, thermophilic lactic acid, and chain-forming (Bacillus spp.) ones. The effect that the microflora has on grass and legume plant productivity, especially when the two kinds of plants are grown together remains understudied, so the main objective of the presented study is to investigate the effect of soil microbiota on the rhizosphere of grass and legume plants and the enhancement of their forage yield after mixed (combined) planting. The obtained results will make it possible not only to forecast the yield but also to reduce the cost of the raw materials used in the production of high-quality forage.
The problem of plants/microbiota interaction is a serious consideration when one selects crops to be grown under certain climatic and environmental conditions of a certain country. According to the study performed by researchers from China Agricultural University combined growth of several crops affects the biodiversity of soil microflora increasing the latter’s quality (Qiao et al., 2012). Studies have demonstrated that the rational use of fertilizers and increased crop rotation within four years can have a direct effect on the phylogenetic structure of soil microflora (Ashworth et al., 2017).
The plants grown simultaneously may affect one another in both positive and negative ways. For instance, indoleacetic acid (IAA) is a plant hormone produced by the leaf tip and buds of young plants. IAA and other auxins facilitate plant growth, dividing and extending the cells (Arkhipova et al., 2005). However, IAA is not plant-specific and is also produced by bacteria (Patten and Glick, 2002). Moreover, multilevel seed-microbe interactions may facilitate shooting (Van der Heijden at al., 2016), which confirms the theory that microbial diversity provides better results than a monomicrobial effect.
On the other hand, seed-microbe interactions can be antagonistic at best. Microbial degradation of seeds, including legume ones, can reduce their productivity (Greenfield, 2000). It may also facilitate soil remineralization (Greenfield, 2000) and improve plant nutrition, but this effect has not been confirmed for many legume crops.
The nutrition conditions and seed combination of mixed planting have their effect on soil fertility and biological activity. The performed studies have demonstrated that using annual legume plants (lupin, vetch, and peas) in a mixed planting with cereal crops increases the biological activity of sod-podzolic, mildly argillaceous soil.
Problem Statement
The gaps in the studies devoted to the effects microflora have on the productivity of the most profitable crops made it essential to study the issue for grass and legume plants. Such studies are of great importance for the preservation and fertility enhancement of the soils of the Kaliningrad Region because they make it possible to obtain well-balanced green forage but also to siderite environment -forming plants in a late-autumn period. That is why, the objective of our study was to determine the effect grass and legume plants had on soil biological activity after spring and summer mixed plantings and how this approach may affect the productive, forage, and sideration aspects of the crop rotation in the Kaliningrad Region.
To achieve the study’s objective, the following tasks had to be resolved:
Methods
The presented study into the productivity of grass and legume mixtures was carried out in 2018-2019 in the experimental field of Kaliningrad Research Institute of Agriculture, a branch of V.R. Williams Federal Research Institute for Fodder Production and Agroecology in the Slavyankoe Settlement of the Polessky District of the Kaliningrad Region.
The soil of the field is sod-podzolic, and moderately cultivated; its composition is mildly argillaceous on moraine loam; weakly gleyic; of average thickness; residual carbonate with low humus content (1.9-2.1). The soil solution reaction is weakly acetous (рН 5.1-5.3). The mobile phosphorus and exchange potassium content changed insignificantly throughout the study varying between 20.5 – 22.2 and 25.0-29.5 mg/g for 100 g of soil, respectively. The seed to be used for planting were pretreated. The pure and mixed grass and legume crops were cultivated following the standard approach using no protecting agents and mineral fertilizers. Before the experiment, the field had been taken by spring triticale as a preceding crop. During the experiment, the following four crops were investigated: millet (Krasava cultivar); blue lupin (Vityaz); field beans (Yantarnye); spring vetch (Yubile’naya 110); field peas (Zaryanka) that were mixed in different proportions (see Table 1).
Table 1. Seeding terms and rates for grass and legume mixtures during the 2018-2019 experiment.
|
Mixture/ seeding term |
Spring planting, 16.04 |
Summer planting, 07.07 |
|||
|
Mixture combination |
Seeding rate in mln of germinating seeds |
Field germination, % |
Seeding rate in mln of germinating seeds |
Field germination, % |
|
|
1 |
Lupun (1.5)+peas (0.3) + beans (0.3) + vetch (0.1) |
2.2 |
96 |
2.2 |
95 |
|
2 |
Lupin (1.5)+peas (0.3)+ beans (0.3)+ vetch (0.1) +millet (2.0) |
4.2 |
96 |
3.2* |
95 |
|
3 |
Lupin (1.5)+millet (2.0) |
3.5 |
96 |
2.5 |
95 |
|
4 |
Lupin (1.5)+peas (0.3)+ beans (0,3)+ vetch (0.1) +millet (2.0) |
4.2 |
96 |
3.2 |
95 |
* The Krasava millet summer seeding rate was reduced to 1 mln of germinating seeds per 1 ha.
The total area of the experimental field of 10800 m2 was subdivided into 12 plots of 900 m2 each. The registration plot area was 100 m2. The four versions of the experiment were repeated 3 times. During the experiment, one performed phenological observations to track the development phases of the spring and summer plants. The plots were weeded out manually, no herbicides were applied. Harvesting and yield accounting included moving the green mass to be weighed per registration plot. The yield of the spring seeding was accounted on July 15, and of the summer seeding – on October 5, 2019.
The assessment methods in the experiment employed microbiological, biochemical, and agrochemical techniques (Moskaleva et al. 2011; Evdokimov, 2018). Analysis of the soil samples employed the following methods: potentiometric analysis to determine рНК; the Kirsanov technique (GOST Р 54650-2011) to estimate the mobile phosphorous and exchange potassium; the Tyurin technique to assess the hummus; the Kappen technique to estimate hydrolytic acidity; the computational method to estimate alkalis saturation; the Sokolov technique to estimate the exchange aluminum. The microbiological activity of the soil was assessed by the team of the laboratory of Chair of Soil Agroecology of Kaliningrad State Technical University supervised by O.M. Bedareva.
The rhizospheric index (RI) of mixed crops was determined using the Dumacheva technique (2012). The index is the ratio of main feeding elements and рНKClboth within and beyond the rhizosphere. To calculate the index the formula RI = А / В was used, where A denotes a feeding element content (or рНKCl) in rhizospheric soil and B - a feeding element content (or рНKCl) in non- rhizospheric soil. To separate rhizospheric and non-rhizospheric soils, mechanical shaking was applied, so the soil that was shaken freely from a root within 5 minutes was regarded as non-rhizospheric, and the soil that got stuck to the root – as rhizospheric.
All the performed analyses were repeated 4 times. The obtained data were processed using Excel following the standard methods and recommendations (Khalafyan, 2007) to be averaged per vegetation period for each of the crop rotations considered.
Results
The performed study has demonstrated that including plants with different linear growth rates in the seeding mixtures makes it possible to set multilayer crops where the bottom layer is taken by lupin and millet and the top one – by beans, vetch, and peas. Such layering allows the plants to use sunlight energy more effectively and prevents soil moisture evaporation. It is noteworthy that during green forage harvesting, the height of the dominating plant (field beans) after summer planting was 164 cm being much higher than that of the beans after spring planting – 94 cm. The same can be said about the heights of all the other plants – they were significantly lower for the spring planting than for the summer one. The lowest height in the experiment was registered for spring lupin (51.8 cm) grown together with millet (71.3 cm)
One of the key factors leading to higher yield in plants is achieving the optimum leaf area concerning environmental conditions. In our experiment, the leaf area was relatively high, reaching its highest for the second (78.9 Km2/ha) and fourth (91.7 Km2/ha) summer seed mixtures (see Table 2).
Table 2. Leaf areas registered in the experiment.
|
Mixture |
Combination |
Leaf area, Km2/ha |
|
|
Spring planting |
Summer planting |
||
|
1 |
Lupin + peas + beans + vetch |
38.9 |
61.2 |
|
2 |
Lupin+ peas + beans + vetch +millet |
43.5 |
78.9 |
|
3 |
Lupin + millet |
31.3 |
52.1 |
|
4 |
Lupin + peas + beans + vetch +millet |
46.4 |
91.7 |
The smallest leaf area was registered for the lupin/millet combination to be 31.1 Km2/ha.
The nutritional value and quality of green forage can not be expressed by a single index for this estimation should take into account such parameters as forage chemical composition; calorie content; digestibility; total, protein, amino acid, mineral, and vitamin nutrition values.
The experiment has demonstrated that the summer mixtures had higher total nitrogen and crude protein contents (2.77–3.47 and 16.41–18.41 %, respectively) due to symbiotic nitrogen fixation activation at optimal temperature and humidity levels. As for the herbage fat and sugar contents, they also maximized in the summer crops and varied from 4.9 to 6.32% for sugar and from 1.28 to 1.45% for fat depending on the seeding rate and seed mixture proportions (see Table 3).
Table 3. Biochemical herbage composition for the 2018-2019 spring/summer seed mixtures.
|
Mixture/ Seeding term |
Biochemical herbage composition, % |
||||||
|
TN |
Phosphorus |
Potassium |
Protein |
Sugar |
Fat |
||
|
1 |
Spring planting |
1.90 |
0.263 |
3.73 |
12.25 |
4.4 |
1.21 |
|
2 |
2.00 |
0.266 |
5.45 |
13.44 |
4.5 |
1.22 |
|
|
3 |
1.98 |
0.248 |
3.01 |
12.10 |
4.0 |
1.20 |
|
|
4 |
2.59 |
0.315 |
5.12 |
15.94 |
4.8 |
1.23 |
|
|
1 |
Summer planting |
2.96 |
0.348 |
6.23 |
16.95 |
5.4 |
1.34 |
|
2 |
3.35 |
0.373 |
6.53 |
17.63 |
5.7 |
1.35 |
|
|
3 |
2.77 |
0.325 |
6.06 |
16.41 |
4.9 |
1.28 |
|
|
4 |
3.47 |
0.395 |
6.77 |
18.41 |
6.2 |
1.45 |
|
The maximum dry mass (28.7 dt/ha) and protein (88.1 kg/ha) yields were obtained from the fourth combination of the seed mixtures. The minimum yields were registered in the third combination – 24.1 dt/ha and 523 kg/ha, respectively (see Table 4).
Table 4. Dry mass yield and nutritional value for the 2018-2019 spring/summer seed mixtures.
|
Mixture/ Seedingterm |
Dry mass yield, dt/ha |
FPU, KU/ha |
Protein output, kg/ha |
Digestibleproteinyield, kg/ha |
FPU yield, KU/ha |
|
|
1 |
Spring planting |
24.9 |
6.65 |
461 |
324 |
21.5 |
|
2 |
25.0 |
6.7 |
629 |
441 |
25.4 |
|
|
3 |
24.1 |
5.56 |
523 |
366 |
21.1 |
|
|
4 |
25.3 |
7.1 |
683 |
478 |
27.4 |
|
|
1 |
Summer planting |
28.2 |
7.9 |
752 |
547 |
33.5 |
|
2 |
27.7 |
8.7 |
773 |
595 |
35.1 |
|
|
3 |
26.8 |
7.3 |
708 |
533 |
28.9 |
|
|
4 |
28.7 |
9.9 |
881 |
637 |
38.7 |
|
|
HCP05 |
1.3 |
|
|
|
|
|
Protein yield from per unit area for the summer crops was on average 1.5 times higher than for the spring ones.
Estimation of the protein and FPU output per 1 ha to assess the quality of the herbage yield has demonstrated that the summer seedings were better by 55.8 % in the first combination, by 38.2% - in the second, by 36.9% - in the third, and by 41.2 % - in the fourth mixture combination. The maximum protein (881 kg/ha) and FPU (38.2 KU/ha) outputs were obtained for the summer mixture of blue lupin, field beans, spring vetch, field peas, and millet.
However, despite the multiple positive results obtained in many research studies, the summer planting of annuals has still not given the place it deserves in the agricultural industry.
More often than not, annual crops are considered a sort of “insurance” to be seeded in case of massive death of spring crops due to improper precipitation or other reasons. In the best-case scenario, these crops are regarded as of secondary importance and recommended to be used as post-cut and cover-ground crops (Khalafyan, 2007).
In soil, nitrogen presents in organic compounds, which makes it inaccessible to plants and limits their growth. Mineralization of such organic compounds that are associated with ammonium release occurs due to ammonifiers. Analysis of the bacteria’s physiological activities has demonstrated that they were very active in the soils taken by grass and legume plants. In the summer crops, their number increased 1.7-2.1 times if compared to the spring ones, which is, probably, due to more favorable environmental conditions when plants are more active and their root exudates contain higher amounts of easily hydrolyzed compounds (see Table 5).
Table 5. Ecologotrophic groups abundance for the 2019 summer seed mixtures.
|
Mixture/ Seedingterm |
Bacteria, mln cells/g a. d. s. |
Actinomycetes, mln cells/g a. d. s. |
Micromycetes, K CFU/g a. d. s. |
||||
|
Ammonifiers |
Aminoautotrophs |
Oligotrophs |
Oligonitrotrophs |
||||
|
1 |
Springplanting |
1.4 |
1.2 |
2.5 |
1.5 |
0.79 |
27.0 |
|
2 |
3.0 |
4.7 |
1.3 |
4.1 |
1.1 |
23.7 |
|
|
3 |
2.5 |
3.8 |
1.7 |
4.5 |
0.88 |
54.1 |
|
|
4 |
3.3 |
4.1 |
2.0 |
4.9 |
1.3 |
27.1 |
|
|
1 |
Summeranting |
1.9 |
2.7 |
2.8 |
2.8 |
0.91 |
22.4 |
|
2 |
4.2 |
5.6 |
2.1 |
9.5 |
1.3 |
25.2 |
|
|
3 |
3.8 |
5.5 |
2.5 |
3.1 |
1.2 |
35.9 |
|
|
4 |
4.1 |
5.7 |
3.1 |
4.2 |
1.4 |
19.8 |
|
|
HCP05 |
0.18 |
0.32 |
0.22 |
0.30 |
0.08 |
2.22 |
|
Among the microflora consuming mineralized nitrogen, bacteria prevail. Their abundance in the media with mineralized nitrogen demonstrates their potential ability to immobilize nitrogen in microbial biomass that increases the amount of biologically bonded nitrogen and reduces unproductive losses. In our experiment, the number of animoautotrophs was highest in the soil taken by wheat, vetch, and peas. Adding small doses of mineral fertilizers activated microbiological processes stimulating the growth of animoautotrophs.
In the summer crops, changing the light regime affected the abundance of oligotrophs in the soil. Prokaryotes in such soils were not abundant due to, probably, sufficient amount of nutriment. The minimum oligotrophic coefficient was registered in the third spring mixture of lupin and millet.
If it comes to hummus, an important role in its formation and decomposition in the soil is played by actinomycetes, whose presence may be a sign of increased mineralization of inaccessible substances. In our experiment, changes in the number of actinomycetes matched the dynamics of nitrogen - mobilizing bacteria that grew in the soil taken by the mixed summer crops. It is also noteworthy that in such soils the proportion of bacteria increased if compared to that of actinomycetes, whose proportion decreased relative to an increasing proportion of legumes.
In the soil taken by the spring mixtures with a high proportion of legumes, the number of micromycetes reduced two times, and 1.5 times on average – in the soils taken by the summer crops if compared to the third combination. The relative soil biogenicity index that reflects the relative bacteria/micromycetes abundance fluctuated significantly varying from 9.4 to 12.8 for the spring crops, and from 12.9 to 18.8 – for the summer mixtures. Changing of the proportion in favor of the bacterial flora evidenced the presence of dehumidified organic matter.
The root exudates of the mixed summer crops had a positive effect on the proliferation of nitrogen-fixing bacteria. The best conditions for such proliferation were provided by the summer crops with a sufficient proportion of legumes, which is, probably, because legumes can assimilate immobile phosphorus compounds.
The mineralization coefficient value of the soils taken by the grass and legume crops was above 1 to be the evidence of intensive nitrogen mobilization in the soil. This is also confirmed by the high microbial transformation rate of plant residues. Its maximum value was registered for the fourth combination of summer mixtures (see Table 6).
Table 6. Microbiological processes intensity in the soils taken by the 2019 spring/summer mixed crops.
|
Mixture/ Seedingterm |
Coefficients/rates |
Relative microflora density, % |
|||||
|
Mineralization |
Oligotrophic |
Oligonitrophic |
Microbialtransformation |
Nitrogen-fixing |
Cellulose-decomposing |
||
|
1 |
Spring planting |
0.90 |
1.87 |
1.13 |
3.23 |
54.8 |
27.6 |
|
2 |
1.60 |
0.45 |
1.36 |
5.06 |
78.4 |
31.0 |
|
|
3 |
1.50 |
0.69 |
1.11 |
4.33 |
37.6 |
22.0 |
|
|
4 |
1.55 |
0.70 |
1.90 |
5.66 |
82.1 |
38.0 |
|
|
1 |
Summer planting |
1.05 |
1.47 |
1.47 |
3.71 |
53.2 |
29.8 |
|
2 |
1.30 |
1.50 |
2.26 |
7.35 |
66.0 |
38.5 |
|
|
3 |
1.50 |
0.66 |
1.41 |
6.43 |
44.1 |
23.0 |
|
|
4 |
1.71 |
1.55 |
2.51 |
7.75 |
79.3 |
53.4 |
|
Oligotrophic bacteria play a significant role in nitrogen mobilization. Their active proliferation in the soils taken by the mixed summer crops can be explained by their low need for nutrients, first hand, nitrogen. Plants consume nitrogen to grow and develop, so oligotrophic bacteria take advantage of it. The high oligotrophic coefficient is evidence that in the soil taken by the mixed summer crops that the last actively consumed nitrogen and its immobilization in the soil increased.
Cellulose decomposition is one of the signs of biological activity in the soil. Both spring and summer mixed crops demonstrated its active decomposition for the combinations with a high proportion of legumes. The abundance of cellulose-decomposing bacteria in the soil taken by the spring crops increased 1.4 times, and 1.7 times – in the soils taken by the summer crops if compared to the third binary combination (lupin and millet).
Analysis of the aggregate soil composition allowed us to see the common patterns soil microflora changed its activity in soil aggregates of different fractions.
Wet sieving demonstrated that aggregate water stability increased together with legume proportion to be the result of processes within the legume rhizosphere and beyond it. When one intensely practices monoculture farming, it results in a significant reduction of agronomically valuable aggregates, especially of mesoaggregates of 2-5 mm in size.
In our experiment, the soil taken by the fourth summer mixture demonstrated the highest microbiological activity. Its microbial cenosis distribution in aggregate fractions of different sizes can be seen in Figure 1.
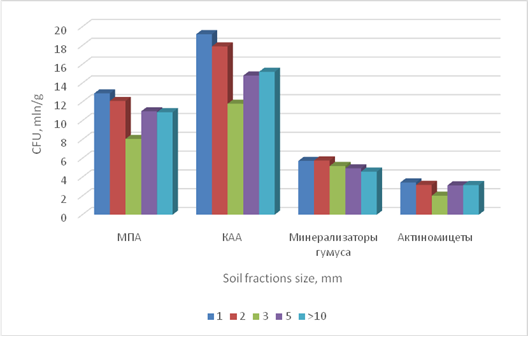
MPA KAA Humus mineralizers Actinomycetes
Figure 1. Microbial cenosis composition of soil fractions different in size.
The activity of beneficial microflora in agronomically valuable 2-5 mm soil particles resulted in increased soil fertility and higher herbage yield in the mixed-grass and legume crops. It was also observed that рНKCl became more acetous increasing the amount of available chemical elements in the rhizosphere. In the summer crops, рНKCl became more alkali increasing the amounts of available nitrogen, phosphorous, and potassium in the rhizosphere. The soil conditions forming in spring complicated the way nutrients reached the rhizosphere of the cereals (millet), which portraited itself in insufficient development of the plant’s elevated part. Opposite processes were observed in the summer crops; they resulted in intensive millet growth and a significant increase of the herbage (see Figure 2).
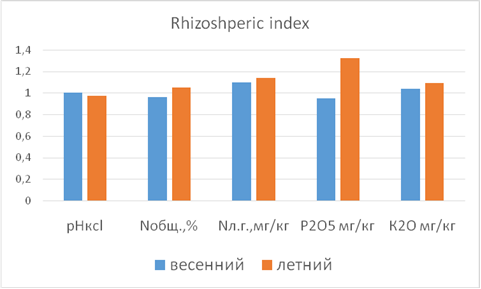
|
pHkcl TN, % Easily hydrolyzed nitrogen mg/kg P2O5 K2O |
|
Spring planting Summer planting |
Figure 2. RI of the mixed spring/summer grass and legume crops.
The performed RI analysis enabled us to clarify the processes occurring within the rhizosphere of legumes and beyond it in the spring/summer mixed crops.
Discussion
We are convinced that in the crop research of the Kaliningrad Region, summer annuals should be given not secondary but a primal role to be covered by dedicated scientific research performed concerning the objectives set by the agricultural industry.
The most perspective way in using summer annuals is the production of high-quality late -autumn green forage. In our experiment, the most abundant in this respect were mixed grass and legume crops of field beans, vetch, millet and lupin, and legume crops of lupin and peas. Their herbage yield in different years depending on soil moisturization reached 80 t/ha.
In the Kaliningrad Region that has favorable conditions in this respect, summer crops can be recommended to be used in crop rotations after harvesting such winter crops as raps and barley or after harvesting winter and spring herbage.
Thus, using spring/summer seed mixtures allows one to obtain high herbage yields per unit area and provide high economic effectiveness and ecological safety while the production of green forage high in protein. Planting summer annuals is an effective method to prevent water and air soil degradation, so it has to be practiced during crop rotations as separately. It also helps exclude excessive soil moisturizing during fall and winter. Autumn sideration with the high-alkaloid lupin varieties planted in summer is the most effective approach to increase the fertility of the soil in the region and allows one to plow under up to 50-80 t/ha of herbage and up to 90 t/ha if root residuals are accounted.
Conclusions
The performed study has shown that including plants with different linear growth rates into seeding material makes it possible to set multilevel crops. In our experiment, the bottom level was taken by lupin and millet, and the top one – by beans, vetch, and peas. This distribution allowed the plants to use sunlight energy more efficiently and prevented moisture evaporation from the soil.
It has also been demonstrated that the leaf area of the mixed crops was significant, reaching its maximum in the second (79.9 K m2/ha) and fourth (91.7 K m2/ha) summer mixtures in combination with blue lupin, filed beans, spring vetch, field peas, and millet. The smallest leaf area was registered in the lupin/millet combination (31.1 K m2/ha)
The tested summer grass and legume mixtures had a high level of total nitrogen and crude protein (2,77–3,47 and 16,41–18,41 %), which was due to symbiotic nitrogen fixation activation at optimum temperature and humidity. As for the herbage fat and sugar contents, they also maximized in the summer crops and varied from 4.9 to 6.32% for sugar and from 1.28 to 1.45% for fat depending on the seeding rate and seed mixture proportions.
The maximum dry grass (27.8 dt/ha) and protein (881 kg/ha) outputs were obtained from the fourth summer mixture and the minimum ones (24,1 dt/ha and 523 kg/ha, respectively) - from a summer mixture of blue lupin and millet. The protein yield in this experiment was 1.5 times higher after the summer planting than after the spring one.
Estimating the herbage yield quality in terms of protein output and FPU has demonstrated that after summer planting the output was higher for all the four mixtures tested (by 55.8, 38.2, 36.9, and 41.2 %, respectively). The maximum protein output (88.1 dt/ha) and FPU (38.7 K units per ha) were obtained after summer planting of a mixture of blue lupin, field beans, spring vetch, field pea, and millet.
It has been verified that the number of ammonifying bacteria after mixed summer planting increased 1.7-2.1 times if compared to that after spring planting, which was due to more favorable conditions when more of the easily hydrolyzed rhizospheric compounds were produced. Actinomycetes are yet another substance that plays an important role in the formation and decomposition of humus and the transformation of other substances. Their presence in studied soil can be an indicator of the increasing mineralization of inaccessible substances in the soil. Our experiments have demonstrated that the highest level of actinomycetes was registered after mixed summer planting and equaled 1.4 mln cells/g a. d. s. Increasing the proportion of legumes in the seeding mixtures led to a reduction in microfungi. Hence, after mixed spring planting, their amount reduced 2 times, 1. 5 times – after mixed summer planting if compared to the third combination (lupin and millet).
The relative soil biogenicity fluctuated significantly, its value changing from 9.4 to 12.8 for the spring and from 12.9 to 18.8 for the summer mixtures.
The mineralization coefficient in the experiment exceeded one to be the evidence of intensive nitrogen mobilization in soil, which is also confirmed by the high coefficient of microbial transformation of plant residuals, whose highest value was registered for the fourth summer mixture.
The performed RI analysis has demonstrated that all the legumes in the experiment triggered similar processes within the rhizosphere and beyond it. In the spring mixtures, they shifted рНKCl to be more acetous increasing the amount of available chemical elements in the rhizosphere, unlike the summer mixture making рНKCl more alkali and increasing the amounts of nitrogen, phosphorous and potassium in the rhizosphere.
References
Arkhipova T.N., Veselov S.U., Melentiev A.I., Martynenko E.V., & Kudoyarova G.R. (2005). Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil, 272, 201–209. https://doi.org/10.1007/s11104-004-5047-x
Ashworth A.J., DeBruyn J.M., Allen F.L., Radosevich M., & Owens P.R. (2017). Microbial community structure is affected by cropping sequences and poultry litter under long-term no-tillage. SoilBiology and Biochemistry, 114, 210-219. https://doi.org/10.1016/j.soilbio.2017.07.019
Benc V.А. (1999). Mixed crops in field forage production of Western Siberia. RASHN, Siberian Branch, SibNII kormov, Novosibirsk, 72 pp (in Russian).
Clune S., Crossin E., & Verghese K. (2017). Systematic review of greenhouse gas emissions for different fresh food categories. J Clean Prod., 140, 766–783.
Dumacheva Е.V. (2012). Soil and rhizosphere interactions in some species of Fabaceae grown in carbonate soils, Fundamental Studies, 9 (part 2), pp 351-355 (in Russian).
Evdokimov I.V. (2018). Methods to detect soil microflora biomass. Russian Journal of Ecosystem Ecology, 3, https://doi.org/10.21685/2500-0578-2018-3-2 (in Russian).
Greenfield L.G. (2000). The microbial decomposition of seeds. Agron Soc New Zeal., 12, 47–51.
Guardia G., Tellez-Rio A., García-Marco S., Martin-Lammerding D., Tenorio J.L., Ibanez M.A., & Vallejo A. (2016). Effect of tillage and crop (cereal versus legume) on greenhouse gas emissions and Global Warming Potential in a non-irrigated Mediterranean field. Agric Ecosyst Environ, 221, 187–197.
Khalafyan А.А. (2007). STATISTIKA 6. Statistical analysis, 3rdedition, Binom-Press, Moscow, 512 pp. (in Russian).
Krasnoperov А.G., & Buyankin N.I. (2019). Spring and summer mixed grass and legume plantings. 2 (30), pp. 144-154 (in Russian).
Moskaleva V.L. (2011). Agroecological role of grass and legume annuals in agricultural biologization. Problems of Agrochemistry and Ecology, 2. 42–45 (in Russian).
Patten C.L. & Glick B.R. (2002). Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl Environ Microbiol, 68(8), 3795-3801. https://doi.org/10.1128/AEM.68.8.3795-3801.2002.
QiaoY.J., Li Z.Z., Wang X., Zhu B., Hu Y.G. & Zeng Z.H. (2012). Effect of legume-cereal mixtures on the diversity of bacterial communities in the rhizosphere. Plant Soil Environ., 58(4), 174–180
Stagnari F., Maggio A., Galieni A., & Pisante M. (2017). Multiple benefits of legumes for agriculture sustainability: an overview. Chem. Biol. Technol. Agric., 4(2). https://doi.org/10.1186/s40538-016-0085-1
Van der Heijden M.G., De Bruin S., Luckerhoff L., Van Logtestijn R.S. & Schlaeppi K. (2016). A widespread plant-fungal-bacterial symbiosis promotes plant biodiversity, plant nutrition and seedling recruitment. ISME J, 10(2), 389-399. https://doi.org/10.1038/ismej.2015.120
Zvyagintsev D.G., Babieva I.P., & Zenova G.M. (2005). Soil biology, Moscow, MSU, 445 pp (in Russian).








