Bioplastic Production from Microalgae and their Applications- A Critical Review
Aluru Ranganadhareddy*,Ponnam Vijetha, Chanda Chandrsekhar
Abstract
Around the world, plastic materials are being produced and consumed more as the global demand for plastic materials increases. Consequently, both marine and terrestrial life have been negatively affected by plastic waste pollution. Multiple health risks are associated with microplastics. The decomposition of plastics takes a long time, and therefore reusing plastics, burning them, chemically treating them, and burying them in landfills are not the best methods to reduce the polluting effects of these plastics. As a result of these characteristics, researchers have searched for alternatives to conventional synthetic plastics that decompose faster. It is easy to collect, process, and use microalgae to make biopolymers. Microalgae are abundant in our ecosystem. There are no known harmful effects associated with microalgae, but they have faster growth rates and are more able to cultivate in wastewater. It is possible to produce biodegradable plastic using polysaccharides in algae. In this study, we examine whether microalgae can produce biodegradable plastics and their economic viability. As a result, two newly identified environmentally friendly approaches are examined in this article: bioplastic production via microalgae, plastic biodegradation, andtheir applications.
Keywords: Bioplastic, Microalgae, Biopolymers, Biodegradation
Introduction
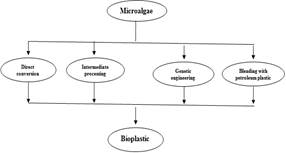
Developing new materials, products, and technologies has become easier due to the growing awareness of environmental sustainability and green chemistry. In recent years, researchers have been interested in developing sustainable polymers from renewable resources because of the long-term persistence of plastics in the environment. Under nutrient-deficient conditions, a wide range of microorganisms produces and accumulate PHAs intracellularly as reserves for carbon and energy. Microorganisms have been observed to spontaneously produce some of these biopolymers in some cases. Biopolymers can be synthesized by fungi, algae, yeast, and bacteria of both gram-positive and gram-negative types (Ranganadha et al., 2019). The annual global production of bioplastic is estimated to be around 1% at present. Plastic production reached 368 million tons in 2020 due to an increase in global demand in recent years (Aman et al., 2022). The current waste management infrastructure is threatened by more than 8 million tons of plastic waste being discarded in the oceans every year (Cywar et al., 2022). The non-degradable nature of synthetic plastics has led to the accumulation of waste on land-filled sites and in the environment. Therefore, several research studies have been conducted on how to create biodegradable plastics from brown, red, and green algae, as they can decompose within a short amount of time (Thiruchelvi et al., 2021). It is possible to produce quality bioplastics from microalgae by using their protein, starch, hemicellulose, lignin, and cellulose, all of which are renewable biomass as shown in Figure 1 (Do Val Siqueira et al., 2021). Due to its ability to decompose more rapidly, biodegradable plastic minimizes pollution. Furthermore, the Earth can be saved from mismanaged plastic waste and the expenditure involved with removing it. Depending on their properties, bioplastics can be classified into two categories: biodegradable or non-biodegradable, and bio-based or fossil-based.
|
|
|
Figure 1. Microalgae conversion to Bioplastic |
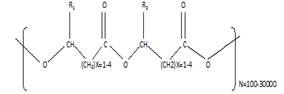
Thermoplastic polymers composed of linear hydroxyalkanoic acid are called PHAs. As one monomeric unit's carboxyl group bonds to another monomeric unit's hydroxyl group, an ester bond is formed (Ranganadha & Chandrasekhar, 2021). Figure 2 shows how they are structured with "n" reaching 35,000 monomers. Monomers of PHA have various R groups from hydrogen atoms to methyl tridecyl (Reddy et al., 2019). The alkyl side chains of phosphoric acid contain usually a saturated alkyl group. However, they can also show a variety of chemical structures that include alkylated, unsaturated, aromatic, branched, epoxidized, and substituted alkyl groups. PHA thermoplastics' side chains can be chemically modified by cross-linking unsaturated bonding (Reddy et al., 2017).
|
|
|
Figure 2. Structure of Polyhydroxyalkanoates with R1 and R2 are alkyl groups (C-C) |
Extraction of PHAs
The recovery of PHAs from microorganisms remains an important stage within the production of such biopolymers as a result it affects their properties; thus, as a result, their industrial applications. PHAs should be retrieved from the cells well once the producing stage since they need to be deposited intracellularly in the living substance of the cells of the microorganisms (cultivation). In this regard, after propagation for the gathering of accumulated PHA, the process of bio-mass may be a crucial stage since it affects the ultimate product qualities (Reddy et al., 2019). Treating, de-stability, cell disruption, recovery, and purification of biopolymers are all processes within the PHA extraction process. Chemical, physical and biological procedures or the combination of these processes may well be employed in these processes to confirm a product of purity with intact physical and thermal properties (Ranganadhareddy, 2022). The first stage within the PHA extraction procedure is to isolate the solid material that is formed of cells containing animate thing biopolymer, from the culture broth, which is typically done by centrifugation. The microorganism plasma membrane also can be broken or destroyed, which may be accomplished through chemical, physical or biological means (Sabathini et al., 2018). A stream of biopolymer, cells containing biopolymer (cells that de-stabilize but don't shatter the cell walls), and cell detritus (mixture of proteins, nucleic acids, lipids, and cell wall fragments) are formed once the cell wall ruptures or de-stabilizes. The biopolymer should then be recovered, which might be done through victimization chemical, biological or physical processes or a mix of those approaches. Once the biopolymer has been recovered by alluviation (centrifugation) or precipitation, the method ends (Ranganadha et al., 2020). The biological approach of harvesting microorganism PHAs could be a difficult procedure that depends on enzymes, as well as lysozymes, nucleases, and proteases, to retrieve the biopolymer. To change cells that accommodate PHA, enzymes are introduced to the culture broth. The light operational conditions, the nice specificity of the enzymes within the reaction of organism cytomembrane proteins while not poignant chemical compound degradation, and therefore the glorious quality of the polymer obtained build this method appealing (Roja et al., 2019; Ranganadha & Chandrasekhar, 2021).
Cultivation of Microalgae
Photosynthetic microorganisms, the microalgae, are gram-negative, eukaryotic, or prokaryotic, colored by photosynthetic pigments, and can colonize any type of environment. As a result of their simple cellular structure, microalgae can quickly develop under adverse conditions, and may thus be an important PHA source, because microorganisms that are in adverse conditions synthesize these polymers (Albuquerque & Malafaia, 2018). As photosynthetic microorganisms, microalgae use inorganic nutrients (nitrogen, phosphorus, carbondioxide, etc. and light energy to produce compounds in the biomass that are valuable. Compared to other heterotrophs like bacteria and yeast, microalgae produce more biomass. This is because heterotrophs must produce their metabolites from organic molecules whereas microbes grow with inorganic compounds and these inorganic compounds are far cheaper than the organic molecules (Anjum et al., 2016). Solar energy can be directly converted into organic molecules through autotrophy (Choi et al., 2020). It is possible to grow microalgae in areas that are inhospitable to agriculture because they can tolerate changes in the environment. Microalgae also have high productivity potential, the microalgae biomass consists of a wide range of bioactive compounds including carbohydrates, proteins, pigments, high value-added molecules like fatty acids, vitamins, antioxidants, polysaccharides, carotenoids, and hydrocolloids, that are useful in various industrial applications (Li &Wilkins, 2020). Microalgae can produce certain compounds by altering the chemical and physical properties of the crops. This means changing the growth conditions can favor the production of PHAs (Mendhulkar & Shetye, 2017).
Factors Influencing the Growth of Microalgae
Several abiotic and biotic factors influence microalgal growth. As well as pathogens like viruses, bacteria, and fungi, other microalgae compete with algae. Culture media contain certain nutrients, pH, light (quality and quantity), salinity, and temperature which constitute the abiotic factors influencing the growth of the microalgae. A PHA is a form of energy storage that microalgae can use to overcome and acclimatize to extreme conditions that are a result of stress factors, both abiotic and biotic. Most of the interference is to and by these stress factors (Dang et al., 2022). Most of the interference to and stimulation of the synthesis of biopolymers comes from abiotic factors (Reddy et al., 2017).
Light intensity and availability influence photosynthesis, therefore directly influencing microalgae growth, biochemistry, and polyhydric acid production. Sunlight reaches the culture medium directly in open culture systems. Due to the low and unequal distribution of light radiation, it can sometimes be observed that growth is limited in these systems (Dang et al., 2022). Alternatively, fluorescent lamps can be used in closed systems utilizing photobioreactors to provide a better distribution of light. The light intensity can also be affected by cell density and pigmentation, which can hinder light penetration and cell growth (Hamad et al., 2018). Due to its influence on metabolic processes and biological reaction rates, temperature directly affects microalgal growth and biomass production. There are different optimal temperatures for different microalgae species (Dang et al., 2022). Microalgae accumulate lipids and PHAs under thermal stress according to several studies (Cruz et al., 2022). In addition to pH, the microalgae growth rate is also affected by the culture medium (Indira et al., 2015). pH 7.0 and 9.0 is the optimal pH range for most species. Because microalgae undergo physiological changes based on the culture medium’s pH, it is crucial to maintain an ideal pH range for crops (Ranganadhareddy et al., 2018; Maghami et al., 2022). There has yet to be a new study done on the relationship between pH changes and PHA production in microalgae cultures. Salinity requirements for microalgae differ based on species. Changes in salinity of the culture medium can adversely affect microalgae growth and composition as a result of osmotic and ionic stress (salt) as well as membrane permeability to ions (Reddy et al., 2019). Salinity changes in the culture medium are primarily caused by losses due to evaporation and precipitation in open systems (AlShehri et al., 2022; Dang et al., 2022). According to Li et al. (2021), it is not directly related to PHA production that salinity stress increases the synthesis of carotenoids and lipids. A microalgae culture medium contains a variety of nutrients, especially carbon, nitrogen, and phosphorus, which are essential to microalgae metabolism, as well as biomass productivity (Cruz et al., 2022). It is believed that inorganic carbon, which is a precursor of photosynthetic reactions, is the main nutrient for autotrophic organisms (Reddy et al., 2019). As nitrogen compounds, such as ammonium, and nitrate are converted to organic matter like enzymes, proteins, and chlorophyll, that have nitrogen, more than 10% of microalgae’s biomass comes from the nitrogen compounds (Ranganadhareddy, 2021; İlhan et al., 2022). Microalgae require phosphorous for their growth, as it is necessary for the synthesis of intermediate compounds, proteins, and lipids. It can be done to increase the formulation of bioproducts like PHAs, by manipulating the content of these nutrients. Increasing the concentration of nitrogen compounds, such as proteins, has been shown to stimulate the build-up of PHAs, carbonaceous compounds, and lipids in some microalgae species (Costa et al., 2018). Microalgal culture media with an insufficient supply of nutrients are often thought to be the main method for stimulating the production of PHAs by microorganisms (Hiroe et al., 2018; Abujamel et al., 2022; Yoong et al., 2022). Abiotic factors in microalgae growth and biochemistry have subsequently been found to be crucial in controlling growth rates, since the production of biomaterials with industrial value, such as polymers, proteins, lipids, and pigments, can be facilitated.
PHAs Production in Microalgae and Biosynthetic Pathways
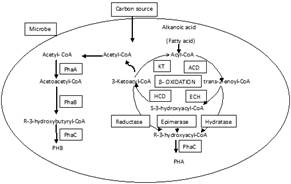
According to numerous studies, PHA synthesis by microalgae is known to occur when microbes grow in a nutrient-deficient condition (Amina et al., 2018; Jallepalli et al., 2022). The most frequent metabolic route, which may be seen in a wide spectrum of bacteria that possess an ability to synthesize PHA and fatty acids, is linked to acetyl CoA, a prevalent precursor. The generation of PHAs by microalgae is thought to follow a similar metabolic mechanism. The PHA is derived from acetyl-coenzyme A (acetyl-CoA) by three enzymatic processes as shown in Figure 3 (Ranganadhareddy, 2022).
|
|
|
Figure 3. Biosynthetic pathway of Polyhydroxtbutyratye in microbes (Ranganadha et al., 2021) |
3- Ketothiolase catalyzes the conversion of two acetyl-CoA molecules as one acetoacetyl-CoA molecule, NADPH-dependent acetoacetyl-CoA reductase catalyzes the reduction of acetoacetyl-CoA to D-3-hydroxybutyryl-CoA and PHA synthase facilitates the association of the D-3-hydroxybutyryl and a PHA molecule through an ester linkage. 3-hydroxy fatty acids are found in β, as well as numerous other PHAs. Between one methyl to thirteen tridecyl carbons make up the pendant group. There are fatty acids having hydroxyl groups at positions 4, 5, and 6 as well as pendent groups carrying substituents or unsaturation, which result in various PHA copolymers and homopolymers (Ravi Teja et al., 2020; Ranganadhareddy & Chandrasekhar, 2021; Daivasigamani et al., 2022).
Applications of Polyhydroxtbutyratye
Agriculture
Bioplastics based on polyhydroxy-alkanoates are commonly used for growing bags, nets, and mulch films in agriculture. Traditional uses of high-density polyethylene (HDPE) included increasing crop quality and yield and protecting crops from birds, insects, and the environment. HDPE-alternative nets are made from bioplastics. The grow bags are made of PHA, which is non-toxic, biodegradable, and eco-friendly (Abd El-malek et al., 2020).
Food Packaging
In today's food industry, packaging challenges are a significant concern, and the industry constantly monitors global requirements and standards. Food industry sustainability and quality standards heavily depend on the development of new bioplastic-based packaging. The use of compostable or degradable bioplastics is vital to meeting the demands of the high-standard storage conditions and the desire for a packaging material that is low in cost, non-polluting, easy to customize, and low in encumbrance. Oxygen permeability, moisture content, and mechanical efficiency are essential attributes of food packaging. Packaging for food must include both water shielding and oxygen shielding (Alexandrovich et al., 2018; Mostafavi & Zaeim, 2020; Anastasova et al., 2022).
Medical Applications
A wide range of uses can be found for polymers in the medical and biological fields (Akhtartavan et al., 2019). The creation of drug delivery systems and tissue engineering devices as biomedical applications was made successful with biodegradable polymers (Daniel et al., 2020). These membranes have holes ranging from 60 to 300 mm in diameter. Nanocelluloses and their composites are heavily used in bioplastic research. Several new developments in nanocellulose-based materials are being investigated in recent years, including 3D printing (Filiz et al., 2021; Mahendiran et al., 2021). Because of their biocompatibility, poly-hydroxy-alkanoates are also used for other medical applications like post-surgical ulcer therapy, wound healing dressings, detection of cancer, heart valves, artificial blood arteries, and bone tissue engineering, etc (Iolanda et al., 2021; Li & Singh, 2021; Behera et al., 2022; Wu et al., 2022).
Conclusion
With microalgal biomass as raw material, bioplastics can be made that solve plastic problems, boost the bioplastic industry, and contribute to the green environment. Our study examines the current state of microalgae bioplastic production. Bioplastics were reviewed for their research, sources, applications, sustainability, and production to clarify the subject. While producing bioplastics with no use of chemicals is ideal, most microalgae biomass must be processed chemically to become bioplastic, resulting in chemical waste. Green technologies can be used to produce these bioplastics. Nevertheless, to improve the time of processing, quality, applications, and costs, subsequent optimization of the downstream process is necessary for producing microalgae bioplastics. Also, adding other biomaterials to microalgal biomass can improve mechanical properties. To manufacture bioplastics and plastic bends, chlorella and spirulina are the most commonly used algae species. Research is needed in microalgae-based bioplastics manufacturing techniques to overcome economic feasibility concerns. As a result of these issues, microalgae-based bioplastics do not find widespread use on the market.
Acknowledgments: None
Conflict of interest: None
Financial support: None
Ethics statement: None
Abd El-malek, F., Aida, F., Sanaa, O., & Heba, K. (2020). Polyhydroxyalkanoates (PHA) from Halomonaspacifica ASL10 and Halomonassalifodiane ASL11 isolated from Mariout salt lakes. International Journal of Biological Macromolecules, 161, 1318-1328. doi:10.1016/j.ijbiomac.2020.07.258.
Abujamel, T. S. (2022). Understanding the mechanisms of bacterial antimicrobial resistance within biofilms. International Journal of Pharmaceutical and Phytopharmacological Research, 12(1), 17-24. doi:10.51847/o5Bt4kEqyT
Akhtartavan, S., Karimi, M., Karimian, K., Azarpira, N., Khatami, M., & Heli, H. (2019). Evaluation of a self-nanoemulsifying docetaxel delivery system. Biomedicine & Pharmacotherapy, 109, 2427-2433. doi:10.1016/j.biopha.2018.11.110.
Albuquerque, P. B., & Malafaia, C. B. (2018). Perspectives on the production, structural characteristics, and potential applications of bioplastics derived from polyhydroxyalkanoates. International Journal of Biological Macromolecules, 107, 615-625.doi:10.1016/j.ijbiomac.2017.09.026.
Alexandrovich, S. O., Yurievna, V. S., Olegovich, P. I., Yurievna, K. N., Ilyinichna, V. L., & Pavlovna, Y. T. (2018). Provision of Microbiological Safety in the Food Industry based on Special Technological Supporting Solutions. International Journal of Pharmaceutical Research & Allied Sciences, 7(1), 103-113.
AlShehri, O. M., Jali, N. M., Almutairi, Y. M., Aljrais, M. M., Alsirhani, A. M., & AlQudairi, A. S. (2022). Common causes of cusp fracture in adults; a systematic review. Annals of Dental Specialty, 10(2), 35-38. doi:10.51847/jBTdUqt7Cx
Aman, S. R., Tushar, C., Chelladurai, H., Tikendranath, V., & Vikas, S. (2022). A comprehensive review on bioplastic production from microalgae. Materials Today: Proceedings, 56, 171-178. doi:10.1016/j.matpr.2022.01.060.
Amina, T., Abess, B., & Yassine, N. (2018). Phenolic content, HPLC analysis and Antioxidant activity extract from Tamarix Articulata. Journal of Advanced Pharmacy and Education Research, 8(4), 1-8.
Anastasova, L., Ivanovska, T. P., Ancevska, A., Petkovska, R., & Petrushevska-Tozi, L. (2022). Applıcatıon of experımental desıgn approach in optımızatıon of qualıty parameters of calcıum- and magnesıum-enrıched mılk. International Journal of Pharmaceutical and Phytopharmacological Research, 12(1), 7-16. doi:10.51847/MtCiwMuW5D
Anjum, A., Zuber, M., Zia, K. M., Noreen, A., Anjum, M. N., & Tabasum, S. (2016). Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: a review of recent advancements. International Journal of Biological Macromolecules, 89, 161-174. doi:10.1016/j.ijbiomac.2016.04.069
Behera, S., Priyadarshanee, M., & Das, S. (2022). Polyhydroxyalkanoates, the bioplastics of microbial origin: Properties, biochemical synthesis, and their applications. Chemosphere, 294, 133723. doi:10.1016/j.chemosphere.2022.133723.
Choi, S. Y., Rhie, M. N., Kim, H. T., Joo, J. C., Cho, I. J., Son, J., Jo, S. Y., Sohn, Y. J., Baritugo, K. A., Pyo, J., et al. (2020). Metabolic engineering for the synthesis of polyesters: a 100-year journey from polyhydroxyalkanoates to non-natural microbial polyesters. Metabolic Engineering, 58, 47-81. doi:10.1016/j.ymben.2019.05.009
Costa, S. S., Miranda, A. L., Andrade, B. B., De Jesus Assis, D., Souza, C. O., de Morais, M. G., Costa, J. A. V., & Druzian, J. I. (2018). Influence of nitrogen on growth, biomass composition, production, and properties of polyhydroxyalkanoates (PHAs) by microalgae. International Journal of Biological Macromolecules, 116, 552-562. doi:10.1016/j.ijbiomac.2018.05.064
Cruz, R. A., Oehmen, A., & Reis, M. A. (2022). The impact of biomass withdrawal strategy on the biomass selection and polyhydroxyalkanoates accumulation of mixed microbial cultures. New Biotechnology, 66, 8-15. doi:10.1016/j.nbt.2021.08.004
Cywar, R. M., Rorrer, N. A., Hoyt, C. B., Beckham, T. G., & Chen, E. Y. X. (2022). Bio-based polymers with performance-advantaged properties. Nature Reviews Materials, 7, 83-103. doi:10.1038/s41578-021-00363-3.
Daivasigamani, S., Chidambaranathan, A. S., & Balasubramanium, M. (2022). A systematic review on the color stability of maxillofacial silicone materials after disinfection and aging procedures. International Journal of Dental Research and Allied Sciences, 2(1), 8-12. doi:10.51847/8qZssQqjrK
Dang, B. T., Bui, X. T., Tran, D. P., Ngo, H. H., Nghiem, L. D., Nguyen, P. T., Nguyen, H. H., Lin, C., Lin, K. Y. A., & Varjani, S. (2022). Current application of algae derivatives for bioplastic production: A review. Bioresource Technology, 347, 126698. doi:10.1016/j.biortech.2022.126698
Daniel, S., Amanda, L., Adriana, L., & Mario, D. (2020). Approaching the environmental problem of microplastics: Importance of WWTP treatments. Science of The Total Environment, 740, 140016. doi:10.1016/j.scitotenv.2020.140016.
Do Val Siqueira, L., Arias, C. I. L. F., Maniglia, B. C., & Tadini, C. C. (2021). Starch-based biodegradable plastics: methods of production, challenges and future perspectives. Current Opinion in Food Science, 38, 122-130. doi:10.1016/j.cofs.2020.10.020.
Filiz, A. K., Joha, Z., & Yulak, F. (2021). Mechanism of anti-cancer effect of β-glucan on SH-SY5Y cell line. Bangladesh Journal of Pharmacology, 16(4), 122-128. doi:10.3329/bjp.v16i4.54872
Hamad, K., Kaseem, M., Ayyoob, M., Joo, J., & Deri, F. (2018). Polylactic acid blends: The future of green, light, and tough. Progress in Polymer Science, 85, 83-127. doi:10.1016/j.progpolymsci.2018.07.001
Hiroe, A., Watanabe, S., Kobayashi, M., Nomura, C. T., & Tsuge, T. (2018). Increased synthesis of poly (3-hydroxydodecanoate) by random mutagenesis of polyhydroxyalkanoate synthase. Applied Microbiology and Biotechnology, 102(18), 7927-7934. doi:10.1007/s00253-018-9230-z
İlhan, N., Telli, S., Temel, B., & Aştı, T. (2022). Investigating the sexual satisfaction mediating role in the relationship between health literacy and self-care of men with diabetes and women's marital satisfaction. Journal of Integrative Nursing and Palliative Care, 3, 19-25. doi:10.51847/sFjL3OLpqg
Indira, M., Venkateswarulu, T. C., Chakravarthy, K., Ranganadha, R. A., & Prabhakar, K. V. (2015). Isolation and Characterization of Bacteriocin Producing Lactic Acid Bacteria from Diary Effluent. Research Journal of Pharmacy and Technology, 8(11), 1560-1565. doi:10.5958/0974-360X.2015.00278.
Iolanda, C., Claudia, P., Rachele, I., Angela, C., Corsaro, M. M., Giovanni, S., & Cinzia, P. (2021). The power of two: An artificial microbial consortium for the conversion of inulin into Polyhydroxyalkanoates. International Journal of Biological Macromolecules, 189, 494-502. doi:10.1016/j.ijbiomac.2021.08.12
Jallepalli, V. R., Thalla, S., Gavini, S. B., Tella, J. D., Kanneganti, S., Yemineni, G., & Nadendla, R. R. (2022). Impact of Patient education on quality of life in gastroesophageal reflux disease. International Journal of Pharmaceutical and Phytopharmacological Research, 12(1), 25-28. doi:10.51847/dAJecTWofD
Li, F., & Singha., A. V. (2021). Recent advancements to enhance the therapeutic efficacy of antiepileptic drugs. Acta Pharmaceutica, 71(4), 527-544. doi:10.2478/acph-2021-0041.
Li, M., & Wilkins, M. R. (2020). Recent advances in polyhydroxyalkanoate production: Feedstocks, strains, and process developments. International Journal of Biological Macromolecules, 156, 691-703. doi:10.1016/j.ijbiomac.2020.04.082
Li, M., Li, K. & Ren, Y. (2021). Nesfatin-1 protects H9c2 cardiomyocytes against cobalt chloride-induced hypoxic injury by modulating the MAPK and Notch1 signaling pathways. Journal of Biological Research-Thessaloniki, 28(1), 1-10. doi:10.1186/s40709-021-00147-4.
Maghami, H., Parhizkar, H., Riasaty, A., Banani, A., Poustfroosh, M., & Hasehmpour-Sadeghian, M. (2022). Identifying the parenting style of students’ parents of an school at Shiraz University of medical sciences. Journal of Advanced Pharmacy Education and Research, 12(1), 91-96. doi:10.51847/tLcoO2yvKI
Mahendiran, B., Muthusamy, S., Sampath, S., Jaisankar, S. N., Popat, K. C., Selvakumar, R., & Krishnakumar, G. S. (2021). Recent trends in natural polysaccharide based bioinks for multiscale 3D printing in tissue regeneration: A review. International Journal of Biological Macromolecules, 183, 564-588. doi:10.1016/j.ijbiomac.2021.04.179.
Mendhulkar, V. D., & Shetye, L. A. (2017). Synthesis of biodegradable polymer polyhydroxyalkanoate (PHA) in cyanobacteria Synechococcus elongates under mixotrophic nitrogen-and phosphate-mediated stress conditions. Industrial Biotechnology, 13(2), 85-93. doi:10.1089/ind.2016.0021
Mostafavi, F. S., & Zaeim, D. (2020). Agar-based edible films for food packaging applications - A review. International Journal of Biological Macromolecules, 159, 1165-1176. doi:10.1016/j.ijbiomac.2020.05.123.
Ranganadha, A. R., & Chandrasekhar, Ch. (2021). Production of polyhydroxybutyrate from marine source- A Review. Indian Journal of Ecology, 48(6), 1829-1836.
Ranganadha, A.R., Sravani, K., Sanjana, N., & Chandrasekhar, Ch. (2021). Production of biopolymer from bacteria – A Review. Environmental and Earth Sciences Research Journal, 8(2), 91-96. doi:10.18280/eesrj.080205
Ranganadha, R. A., Krupanidhi, S., Venkateswarulu, T. C., Bharath, K. R., Sudhakar, P., & Vidyaprabhakar, K. (2019). Molecular characterization of a biopolymer producing bacterium isolated from sewage sample. Current Trends in Biotechnology and Pharmacy, 13(3), 325-335.
Ranganadha, R. A., Vidyaprabhakar, K., Venkateswarulu, T. C., Krupanidhi, S., Nazneen Bobby, Md., Abraham, P. K., Sudhakar, P., & Vijetha, P. (2020). Statistical optimization of Polyhydroxtbutyratye (PHB) production by novel Acinetobacternosocomialis RR20 strain using Response Surface Methodology. Current Trends in Biotechnology and Pharmacy, 14(1), 62-69.
Ranganadhareddy, A. (2021). Screening and biochemical characterization of PHB producing bacterium isolated from marine sample. Indian Journal of Ecology, 48(4), 1169-1172.
Ranganadhareddy, A. (2022). A Review on Production of Polyhydroxyalkanoates in Microorganisms. Journal of Biochemical Technology, 13(1), 1-6. doi:10.51847/Uo3EEbmgID
Ranganadhareddy, A., Venkateswarulu, T. C., Sudhakar, P., Krupanidhi, S., & Vidyaprabhakar, K. (2018). Optimization of process parameters for Polyhydroxy butyrate production from isolated Acinetobacternosocomialis RR20 through submerged fermentation. Current Trends in Biotechnology and Pharmacy, 12(2), 16-25.
Ravi Teja, K. S., Abraham Peele, K., Venkateswarulu, T. C., Vijetha, P., Krupanidhi, S., & Ranganadha, R. A. (2020). Computational Modeling of Chitinase (Cs CHIO) Protein from Chitiniphilusshinanonensis. Indian Journal of Ecology, 47(11), 151-153.
Reddy, A. R., Kumar, R. B., & Prabhakar, K. V. (2017). Isolation and Identification of PolyHydroxyButyrate (PHB) producing bacteria from Sewage sample. Research Journal of Pharmacy and Technology, 10(4), 1065-1069. doi:10.5958/0974-360X.2017.00193.7
Reddy, A. R., Peele, K. A., Krupanidhi, S., Prabhakar, K. V., & Venkateswarulu, T. C. (2019). Production of Polyhydroxtbutyratye from AcinetobacternosocomialisRR20 strain using modified mineral salt medium: a statistical approach. International Journal of Environment Science and Technology, 16(10), 6447-6452. doi:10.1007/s13762-018-2102-3
Roja, K., Ruben, S. D., Susaimanickam, A., & Thangavel, M. (2019). Extraction and characterization of polyhydroxyalkanoates from marine green alga and cyanobacteria. Biocatalysis and Agricultural Biotechnology, 22, 101358. doi:10.1016/j.bcab.2019.101358.
Sabathini, H. A., Windiani, L., Dianursanti, Gozan, M., Kusrini, E., Juwono, F. H., Yatim, A., & Setiawan, E. A. (2018). Mechanical Physicial Properties of Chlorella-PVA-based Bioplastic with Ultrasonic Homogenizer. E3S Web of Conferences, 67, 03046. doi:10.1051/e3sconf/20186703046.
Thiruchelvi, R., Aryaman, D., & Sikdar, E. (2021). Bioplastics as better alternative to petro plastic. Materials Today: Proceedings, 37, 1634-1639. doi:10.1016/j.matpr.2020.07.176
Wu, N., Tu, Y., Fan, G., Ding, J., Luo, J., Wang, W., Zhang, C., Yuan, C., Zhang, H., Chen, P., et al. (2022). Enhanced photodynamic therapy/photothermo therapy for nasopharyngeal carcinoma via a tumour microenvironment-responsive self-oxygenated drug delivery system. Asian Journal of Pharmaceutical Sciences, 17(2), 253-267. doi:10.1016/j.ajps.2022.01.002
Yoong, S. Q., Wang, W., Seah, A. C. W., Kumar, N., Gan, J. O. N., Schmidt, L. T., Lin, Y., & Zhang, H. (2022). Study of the self-care status and factors related to it in heart failure patients. Journal of Integrative Nursing and Palliative Care, 3, 31-35. doi:10.51847/Lqz1ms7fB8