A Review on Production of Polyhydroxyalkanoates in Microorganisms
Aluru Ranganadhareddy*
Abstract
Plastic waste is a concern that is growing due to its nondegradable nature. Many species of microorganisms naturally produce polyhydroxyalkanoates, macromolecule-polyesters that may be used to replace conventional plastics. A variety of microorganisms can completely biodegrade PHAs within a year, which is unlike petroleum-derived plastic that takes decades to degrade. Carbon dioxide and water are produced during this biodegradation, which is returned to the environment. Many methods of mass-producing PHAs have been attempted. Genetically modified microorganisms can be used to establish novel production mechanisms. PHA production, as well as the expression of a few genes, must be maximized in the host for this task. Despite significant advances that have been achieved in generating transgenic organisms, acquiring large quantities of PHA at reasonable costs continues to remain a difficulty. The increasing awareness as well as the promising nature of utilizing microorganisms as a source of polyhydroxyalkanoates are highlighted in this review.
Keywords: Polyhydroxyalkanoate, Microorganism, Bioplastic, Polyhydroxtbutyratye
Introduction
Our planet has been plagued by a massive accumulation of non-degradable wastes, due to the growth of the human population. As far as the environment is concerned, plastic waste has become a major issue (Padervand et al., 2020; Alanazi et al., 2022). In addition to taking decades for conventional plastics to decompose in nature, they may also generate toxins in the process of degradation. There is therefore a strong interest in producing plastics in a manner that is "environmentally friendly" by using recycled materials (Bilhalva et al., 2018). Also contributing to the popularity of bioplastics is the declining availability of petrochemicals. Petroleum products have been widely employed as a major energy source in industrial processes and as building materials in advanced economies. Non-renewable resources, on the other hand, are scarce, and the latest work predicts that, by 2025, consuming fossil fuels will surpass discovering them, given the rate of discovery and previous fuel usage trends. Because our economy is still heavily dependent on oil, this is a global issue. Plastic is currently consumed in the world in an amount of 140 million tons per year. The production of these plastics consumes about 150 million tonnes of carbon fuels. The characteristics of all structural materials made of carbon are determined by long sequences of carbon-carbon bonds (e.g., plastics, foams, coating, and adhesives). The world's problem is to see if we can replace the non-sustainable source of these lengthy carbon arrays with a sustainable and renewable supply.
Bioplastics, unlike plastics made from petroleum, are made up of natural biopolymers that are manufactured and catabolized by a variety of species and do not harm the host organism (Reddy et al., 2019). As a result of stress, these polymers accumulate in microbial cells as storage materials (Bátori et al., 2018). Polyhydroxyalkanoates (PHAs) and their derivatives are the most widely used microbial bioplastics (Kawamura et al., 2021). A lucent granule of PHA was first observed in bacterial cells by Beijerinck in 1888 (Choi et al., 2020). PHAs were first reported by Lemoigne as an unknown compound in the form of polyhydroxybutyrate (PHB), a home polyester comprising the three hydroxybutyric acids (Sathya et al., 2018). In the subsequent 30 years, there was little interest in this unknown material. By 1958, Macrae and Wilkinson had published their first report on the PHB's functions (Bhatia et al., 2019; Hoang et al., 2022). They discovered that PHB generated by Bacillus cereus and Bacillus megaterium biodegrades quickly. PHB's popularity grew dramatically from this point forward. Researchers began experimenting with PHBs and other PHAs in the years that followed and realized how these biopolymers were useful (Zia et al., 2017). An overview of PHAs' chemical structure and properties is presented in the current review. After that, microorganisms are examined for their synthesis of PHA. Last but not least, several ways for synthesizing PHA in plants using genes encoding PHA synthases are demonstrated. The market for PHA polymers has been discussed in detail.
The Physical Properties of Phas and Their Monomer Makeup
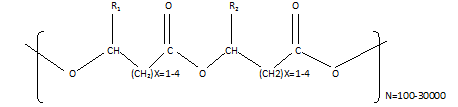
Poly-hydroxyalkanes comprise 3-hydroxy fatty acid-based linear head-to-tail polymers (Figure 1). In general, PHA has 103 to104monomers gathered into inclusions with a diameter of up to 0.5 microns, with accumulating monomers accumulating as inclusions. Both Gram-negative and Gram-positive bacteria can produce and store these inclusions without harming the hosts (Ravi Teja et al., 2020). A nutrient imbalance can lead to PH accumulation when cells receive too much carbon with insufficient nitrogen, phosphorus, and oxygen (Reddy et al., 2019). Insoluble biopolymers are formed from soluble molecules by the bacteria to store excess nutrients. When normal growth conditions return, biopolymers become mobilized.
|
|
|
Figure 1. Structure of Polyhydroxyalkanoates with R1 and R2 are alkyl groups (C-C) |
Depending on the organism's physio-chemical characteristics the composition, number, and size of granules may change. (Buzarovska et al., 2018). The number of carbons in alkyl groups, which take up the R configuration at the C-3 position in all characterized PHAs, can range from one (C1) to more than 14 carbons (C14). PHAs can be categorized into three types based on the size of their monomers. PHAs with up to C5 monomers are known as short-chain length PHAs (scl- PHA). mcl-PHAs and lcl-PHAs are PHAs that include C6–C14 and N C14 monomers, respectively. Mcl-PHAs are considered rubbers and elastomers, but scl-PHAs are akin to traditional plastics (Raberg et al., 2018). As an example of functional modification of the monomers, unsaturated and halogenated branched chains have been introduced into the bioplastic to improve its properties. Additionally, heteropolymers can be produced through polymerization between different kinds of monomers. PHB, the most prevalent kind of scl-PHA, is the most thoroughly researched 3-hydroxybutyric acid homopolymer. PHA polymers including 3-hydroxybutyrate (3HB), 3-hydroxyvalerate (3HV), and 3-hydroxyhexanoate (3HH) monomers can be made in a variety of ways. The majority of bacteria produce either scl-PHAs with predominantly 3HB units or mcl-PHAs with 3-hydroxyoctanoate (HO) and 3-hydroxydecanoate (HD) as primary monomers (Ryan & Walsh, 2016). It has been reported that bacteria synthesize more than 150 different types of PHAs (Wu et al., 2016). PHAs derived bacterial cells have characteristics analogous to that of conventional polymers such as polypropylene (Hungund et al., 2018). Many microorganisms can degrade PHAs quickly (3–9 months) by using their own PHA depolymerases to produce carbon dioxide and water (Jendrossek 2020). Biodegradable materials, recyclable and considered natural materials, are available from renewable resources. This makes PHAs an ideal substitute for petrochemical thermoplastics (Iordanskii et al., 2017; Aldhairyan et al., 2022). The broad variety of monomers found in PHAs can create an array of physical characteristics. Homopolymer PHBs are stiff and brittle bioplastics, which have limited applications. In addition, mcl-PHAs can also be modified to modify rubber properties. Elastomers and sticky materials are often formed of PHAs with longer monomers. PHA copolymers largely composed of HB with a small percentage of longer chain monomers such as HO, HV, or HH are flexible and robust. In addition to food containers, they can also be used in bottles, razors, and packaging materials (Alexandrovich et al.,2018). Paper, film, and cardboard can be treated with PHA latex to create a water-resistant layer (Ravi Teja et al., 2020). PHB and co-polymer P(HB-HV) were utilized as a water-resistant coating behind the diaper sheets in the United States (Gajjar & King, 2014; Nurcahyo et al., 2022). ICI/Zeneca and Monsanto marketed this copolymer P(HB–HV) for its fluidity and wear resistance under the brand name Biopol TM until 1995. PHAs are also employed in the production of fiber products, such as nonwoven textiles. Long-chain hydroxyacid PHAs have been utilized in pressure-sensitive adhesive compositions (Requena et al., 2020). Many PHAs are also biocompatible in addition to being biodegradable. They are broken down into 3-hydroxyacids, which are found in mammals naturally. Gauzes, implants, suture, osteosynthetic materials, filaments, as well as matrix materials for a gradual release of medicines and in vitro cell cultures are among the medical applications of these PHAs (Anjum et al., 2016; Chen& Zhang, 2018; Shaheen et al., 2022).
PHA synthesis in Microorganisms
Genes and enzymes involved in PHA synthesis
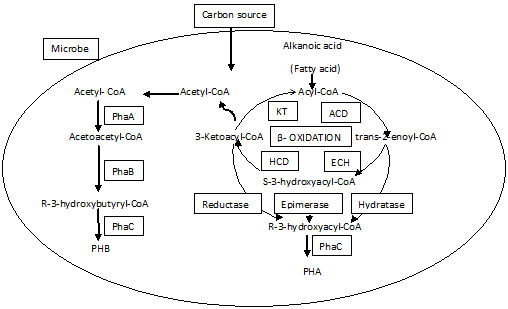
The Halobactericeae family of the Archea encompasses several species of bacteria that possess the ability to produce PHAs. It was found that there are nearly 300 species of such PHA-producing microorganisms exist that add up to the ever-growing list (Morya et al., 2018). Although there are numerous chemical forms of PHAs present, the most eminent and widely synthesized form is PHB (Das et al., 2018). PHB production is one of the most uncomplicated biosynthetic pathways (Gao et al., 2018). The action of three enzymes and their associated transcribing genes is required for the synthesis of PHB (Figure 2) (Zhao et al., 2019). The fusion of two acetyl-CoA molecules to generate acetoacetyl-CoA is the first step in the process, and it is catalyzed by the enzyme -ketothiolase, which is encoded by the phaA gene. The acetoacetyl-CoA formed in the earlier step is now reduced into (R)-3-hydroxybutyryl-CoA with the aid of an NADPH-reliant enzyme acetoacetyl-CoAreductase (Sharma & Dhingra, 2021). The phaB gene is responsible for the encoding of the acetoacetyl-CoAreductase enzyme. The polymerization of (R)-3-hydroxybutyryl-CoA monomers occurs in the last stage, which is catalyzed by the PHA synthase enzyme expressed by the phaC gene (Hiroe et al., 2018).
|
|
|
Figure 2. Biosynthetic pathway of Polyhydroxtbutyratye in microbes (Ranganadhareddy& Chandrasekhar, 2021) |
Ralstonia eutropha, earlier known as Alcaligenes eutrophus, comprises a PHA synthase that interacts with a limited set of substrates with chain lengths ranging from C3 to C5, with a preference for C4 substrates (Brigham, 2019). Hence, the PHA end product synthesized by this process is monomers with a short chain length. All of the enzymes involved in PHB formation are found in the cytosol of the cell, and PHB accumulation is likewise found in the same region within the cell (Mannina et al., 2020). Bacteria can produce a variety of PHAs, and this synthesis is not just limited to PHB (Skariyachan et al., 2018). Many bacteria, including Ralstonia eutropha, have been able to produce various PHAs with C3 to C5 monomers as a result of changes in the amount and source of carbon in the nutrient broth. The addition of propionic or valeric acid in glucose medium, for example, results in the creation of a copolymer with HB and HV [P(HB-HV)]. The condensation of propionyl-CoA with acetyl-CoA is catalysed by ketothiolase in this process (3-ketothiolase, bktB). The reduction of 3-ketovaleryl-CoA to (R)-3-hydroxyvaleryl-CoA and subsequent polymerization to form P(HB-HV) are then subjected to similar enzymes which participate in the manufacture of PHB. Acetoacetyl-CoA reductase and PHA synthase are enzymes that are involved in both stages and PHB synthesis (Ranganadhareddy & Chandrasekhar, 2021). A variety of hydroxyacyl-CoAthio-esters are used as substrates by PHA synthases extracted from numerous bacterial species. The PHA synthase classification is done based on the content in their subunit, and they are segregated into four classes on this basis. The genes that encode the enzymes that are vital for the synthesis of PHA are cloned from the source of a natural origin of the polymer. Anti-thiolase antibodies were used to clone the phaA gene from Zoogloea ramigera for the first time (Sharma & Dhingra, 2021). The presence of the phaB gene in Zoogloea ramigera, Paeacoccus denitrificans, and Rhizobium meliloti was discovered to be in the same operon, but phaC was in a separate operon (Miao et al., 2021). In Acinetobacter spp., Pseudomonas acidophila, Ralstonia eutropha, and Alcaligenes latus, a phaCAB operon is formed by the pha genes of these microorganisms even though the genes do not share the same sequence in these species. The genomes might carry numerous copies of operons in some situations. There is a probability of the PHA synthase having two sub-units (PhaC and PhaE) and this situation holds true in the case of Chromatium vinosum, Thiocap sap fennigi, Thiocystis violacea, and Synechocystis sp. PCC 6803. The function of this type-3 synthase includes the catalysis of the production of scl-PHAs, and the polymerization of scl- and mcl-monomers (Katayama et al., 2018).
Not only have these fundamental enzymes but there existed few other enzymes that enable PHA synthesis indirectly. The phaJ gene in Aeromonas caviae is responsible for encoding the enoyl-CoA flanks PHA synthase gene(phaC). Enoyl-CoAhydratase plays a role as a catalyst in the (R)-specific hydration of 2-enoyl-CoA which in turn supplies (R)-3-hydroxyacyl-CoA monomers for the synthesis of PHA by fatty acid β-oxidation pathway (Zhang et al., 2019). P.oleovorans, P. aeruginosa, Burkholderia caryophylli can synthesize mcl-PHAs while, R. eutropha is unable to do so(Pérez-Nava et al., 2021). These organisms accommodate two phaC genes which are split apart by a phaZ gene in the phaC1ZC2D operon. These genes encode for an enzyme called PHA depolymerase, whose significance remains mysterious, even though it somehow contributes to the PHA synthesis (Mozejko-Ciesielska et al., 2019). P. aeruginosa and P. oleovorans utilize the fattyacid-oxidation pathway's intermediates directly, resulting in large molecules of 3-hydroxyacyl CoA (Raza et al., 2018; Zibi et al., 2022). The P. oleovorans contains PHA synthase which enables the polymerization of monomers leading to the formation of polymers with higher molecular weight and better elastic properties. An extra cluster (phaFl) is present in many microorganisms, and the location of this cluster is downstream from the phaC1ZC2D operon. The role of Phal is the stabilization and formation of granules. And the function of PhaF is to regulate and stabilize the granules (Lim et al., 2018).
Production of PHA copolymers
Stiff crystalline materials that are brittle were formed by scl-PHA homopolymers (C3-C5) such as PHB. The limited flexibility of these scl-PHA homopolymers results in a restricted number of applications. The PHB homopolymer consists of a C4 monomer as its only component tends to degrade at temperatures slightly beyond the melting point, making the homopolymer difficult to handle (Ryan& Walsh, 2016). Unlike the PHB homopolymer made of C4 monomer, the polymers that are built only with mcl-PHA tend to be semi-crystalline thermoplastic elastomers. This makes the mcl-PHA containing polymer possess mechanical properties improved by reinforcement. Based on the mol percent composition of subunits that make up the polymer, the scl-mcl-copolymers preserve better material characteristics than polymers having either mcl- or scl- monomers (Snoch et al., 2019). The majority of the scl-mcl copolymers are made up of c4 monomers with a minimal count of c6 monomers and exhibit the property that is identical to polypropylene. Flexible and tough material can be obtained from the scl-mcl-PHA copolymer of HB and HH [P(HB-HH)]. This HB and HH [P(HB-HH)] copolymer comprise many desirable qualities such as improved flexibility, reduced crystallinity, melting point, simple processing, and increased strength. Many laboratories tried to reap the benefits of these enhanced properties by trying to manufacture a few selective mcl-PHA copolymers in bacteria. The synthesis of mcl-PHA and copolymers is initiated by inserting pha genes from several biological compounds into E. coli, such as the phaC gene from Pseudomonas sp. The phaC1 gene from P. oleovorans in fadA and fadB strains collected mcl-PHAs when cultured on C8 to C18 fatty acids, with an increase in yield attained by using inducible promoters. By inserting the phaC1 and phaC2 genes from Burkholderia caryophylli and P. aeruginosa into the recombinant fadB mutant of E. coli, PHA copolymers including HH, HD, and HO were synthesized. The hbcT gene from Clostridium kluyveri, with a function of encoding 4-hydroxybutyric acid-CoAtransferase, and phaC from R. eutropha were co-transformed with E. coli cells (Gutiérrez-Gómez et al., 2019). The bacteria when supplied with 4HB during its growth resulted in the accumulation of P(4HB) homopolymer, constituting up to 20% of the cell dwt of the cell. The presence of glucose plays a major role in the type of polymer that will be produced, while its presence results in the production of P(4HB) homopolymer, whereas its deficiency leads to the synthesis of P(HB-4HB) copolymer despite the absence of phaA and phaB genes (Najah et al., 2019). Varying the concentration levels of fatty acid and glucose in the medium proven to affect the monomer makeup of PHA copolymer. Through the selection of specific bacterial strains, 43% cell dwt P(HB-HV) copolymer synthesis was obtained in modified E. coli (Lim et al., 2018). To maximize the copolymer synthesis, various fermentation and feeding procedures were implemented. Because E. coli is not a natural PHA producer, boosting its multiplication was challenging.
Conclusion
A substitute to petroleum-based plastics is a substance stored by microorganisms that is biodegradable which is known as PHA. For the exploration of various PHB accumulating strains, the environment can be excavated. By displacing non-biodegradable inorganic plastics, Polyhydroxtbutyratye contribute to the creation of a sustainable environment. To compete in a market driven by thermoplastics, it is important to produce cost effective biodegradable polymers. By reducing the production cost of biodegradable polymers their application range can be broadened. The imperative properties of biopolymers like biodegradability and biocompatibility have made them compatible for usage in packaging, biomedical field, aquaculture field and antifouling. Therefore, with the help of this report, it can be said that PHB is a resilient class of biopolymers that can be used for better designing of new applications in various fields.
Acknowledgments: None
Conflict of interest: None
Financial support: None
Ethics statement: None
Alanazi, A. A., Wajdi, F. A., Al Issa, M. S., Fallatah, A. A., Shaker, A. O., AlHatim, A. A., Alqubali, M. K., Alshammari, R. H., Alghasham, B. A., Almohammedali, H. Y., et al. (2022). An Overview on Klinefelter’s: Clinical Features and Management in Pediatric Population. International Journal of Pharmaceutical Research and Allied Sciences, 11(1), 1-5. doi:10.51847/GrHPjQ9TYY
Aldhairyan, A. H., Alyami, S. S. H., Alsaad, A. M. S., Al Shuqayfah, N. I., Alotaibi, N. A., Mujammami, N. M., Alkhathami, J. F., AlZahrani, Y. A., Ashaari, A. Y., & Alshehri, M. A. (2022). Gastroesophageal Reflux Disease: Diagnosis and Management Approach, Literature Review. World Journal of Environmental Biosciences, 11(1), 1-3. doi:10.51847/EvuxMWxAai
Alexandrovich, S. O., Yurievna, V. S., Olegovich, P. I., Yurievna, K. N., Ilyinichna, V. L., & Pavlovna, Y. T. (2018). Provision of Microbiological Safety in The Food Industry Based on Special Technological Supporting Solutions. International Journal of Pharmaceutical Research & Allied Sciences, 7(1), 103-113.
Anjum, A., Zuber, M., Zia, K. M., Noreen, A., Anjum, M. N., & Tabasum, S. (2016). Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: a review of recent advancements. International Journal of Biological Macromolecules, 89, 161-174. doi:10.1016/j.ijbiomac.2016.04.069
Bátori, V., Åkesson, D., Zamani, A., Taherzadeh, M. J., & Horváth, I. S. (2018). Anaerobic degradation of bioplastics: A review. Waste Management, 80, 406-413.
Bhatia, S.K., Gurav, R., Choi, T.R., Jung, H.R., Yang, S.Y., Song, H.S., Jeon, J.M., Kim, J.S., Lee, Y. K., & Yang, Y.H. (2019). Poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) production from engineered Ralstonia eutropha using synthetic and anaerobically digested food waste derived volatile fatty acids. International Journal of Biological Macromolecules, 133, 1-10.
Bilhalva, A. F., Finger, I. S., Pereira, R. A., Corrêa, M. N., & Burkert Del Pino, F. A. (2018). Utilization of biodegradable polymers in veterinary science and routes of administration: a literature review. Journal of Applied Animal Research, 46(1), 643-649. doi:10.1080/09712119.2017.1378104
Brigham, C. (2019). Perspectives for the biotechnological production of biofuels from CO2 and H2 using Ralstonia eutropha and other ‘Knallgas’ bacteria. Applied Microbiology and Biotechnology, 103(5), 2113-2120. doi:10.1007/s00253-019-09636-y
Buzarovska, A., Dinescu, S., Chitoiu, L., & Costache, M. (2018). Porous poly (L-lactic acid) nanocomposite scaffolds with functionalized TiO2 nanoparticles: Properties, cytocompatibility and drug release capability. Journal of Materials Science, 53(16), 11151-11166. doi:10.1007/s10853-018-2415-0
Chen, G. Q., & Zhang, J. (2018). Microbial polyhydroxyalkanoates as medical implant biomaterials. Artificial Cells, Nanomedicine, and Biotechnology, 46(1),1-18. doi:10.1080/21691401.2017.1371185
Choi, S.Y., Rhie, M.N., Kim, H.T., Joo, J.C., Cho, I.J., Son, J., Jo, S.Y., Sohn, Y.J., Baritugo, K. A., Pyo, J., et al. (2020). Metabolic engineering for the synthesis of polyesters: a 100-year journey from polyhydroxyalkanoates to non-natural microbial polyesters. Metabolic Engineering, 58, 47-81.
Das, S., Majumder, A., Shukla, V., Suhazsini, P., & Radha, P. (2018). Biosynthesis of Poly (3-hydroxybutyrate) from Cheese Whey by Bacillus megaterium NCIM 5472. Journal of Polymers and the Environment, 26(11), 4176-4187. doi:10.1007/s10924-018-1288-2
Gajjar, C. R., & King, M. W. (2014). Current Applications of Biotextiles and Future Trends. Resorbable Fiber-Forming Polymers for Biotextile Applications, 67-75.
Gao, R., Zhang, R., Zhang, C., Liang, Y., & Tang, W. (2018). LncRNA LOXL1-AS1 promotes the proliferation and metastasis of medulloblastoma by activating the PI3K/AKT pathway. Analytical Cellular Pathology, 2018. doi:10.1155/2018/9275685
Gutiérrez-Gómez, U., Servín-González, L., & Soberón-Chávez, G. (2019). Role of β-oxidation and de novo fatty acid synthesis in the production of rhamnolipids and polyhydroxyalkanoates by Pseudomonas aeruginosa. Applied Microbiology and Biotechnology, 103(9), 3753-3760. doi:10.1007/s00253-019-09734-x
Hiroe, A., Watanabe, S., Kobayashi, M., Nomura, C. T., & Tsuge, T. (2018). Increased synthesis of poly (3-hydroxydodecanoate) by random mutagenesis of polyhydroxyalkanoate synthase. Applied Microbiology and Biotechnology, 102(18), 7927-7934. doi:10.1007/s00253-018-9230-z
Hoang, T. T. V., Nguyen, T. H., Nguyen, T. T. T., Hoang, L. P. T., Ho, T. T. T., Nguyen, T. H. T., & Nguyen, T. T. M. (2022). Research Factors Affecting Students’ Academic Results in Learning Project Subjects Oriented CDIO In Vinh University. Journal of Organizational Behavior Research, 7(1), 14-28. doi:10.51847/SntPtYuASo
Hungund, B. S., Umloti, S. G., Upadhyaya, K. P., Manjanna, J., Yallappa, S., & Ayachit, N. H. (2018). Development and characterization of polyhydroxybutyratebiocomposites and their application in the removal of heavy metals. Materials Today, 5(10), 21023-21029.
Iordanskii, A., Ol’khov, A., Karpova, S., Kucherenko, E., Kosenko, R.Y., Rogovina, S., Chalykh, A., & Berlin, A. (2017). Influence of the structure and morphology of ultrathin poly (3-hydroxybutyrate) fibers on the diffusion kinetics and transport of drugs. Polymer Science, Series A, 59(3), 352-362. doi:10.1134/S0965545X17030075
Jendrossek, D. (Ed.). (2020). Bacterial Organelles and Organelle-like Inclusions (pp. 243-275). Springer International Publishing.
Katayama, N., Iijima, H., & Osanai, T. (2018). Production of bioplastic compounds by genetically manipulated and metabolic engineered cyanobacteria. Synthetic Biology of Cyanobacteria, 155-169.
Kawamura, Y., Mori, K., & Amachi, S. (2021). Reductive deiodination of 2, 4, 6-triiodophenol by Vallitalea sp. strain TIP-1 isolated from the marine sponge. Journal of Bioscience and Bioengineering, 132(2), 154-160.
Lim, J. H., Rhie, H. G., & Kim, J. N. (2018). Identification and Analysis of Putative Polyhydroxyalkanoate Synthase (PhaC) in Pseudomonas fluorescens. Journal of Microbiology and Biotechnology, 28(7), 1133-1140. doi:10.4014/jmb.1803.03006
Mannina, G., Presti, D., Montiel-Jarillo, G., Carrera, J., & Suárez-Ojeda, M. E. (2020). Recovery of polyhydroxyalkanoates (PHAs) from wastewater: A review. Bioresource Technology, 297,122478. doi:10.1016/j.biortech.2019.122478
Miao, C., Meng, D., Liu, Y., Wang, F., Chen, L., Huang, Z., Fan, X., Gu, P., & Li, Q. (2021). Biosynthesis of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) in metabolically recombinant Escherichia coli. International Journal of Biological Macromolecules, 193, 956-964. doi:10.1016/j.ijbiomac.2021.10.183
Morya, R., Kumar, M., & Thakur, I. S. (2018). Utilization of glycerol by Bacillus sp. ISTVK1 for production and characterization of polyhydroxyvalerate. Bioresource Technology Reports, 2, 1-6.
Mozejko-Ciesielska, J., Szacherska, K., & Marciniak, P. (2019). Pseudomonas species as producers of eco-friendly polyhydroxyalkanoates. Journal of Polymers and the Environment, 27(6), 1151-1166. doi:10.1007/s10924-019-01422-1
Najah, S., Saulnier, C., Pernodet, J. L., & Bury-Moné, S. (2019). Design of a generic CRISPR-Cas9 approach using the same sgRNA to perform gene editing at distinct loci. BMC Biotechnology, 19(1), 1-8. doi:10.1186/s12896-019-0509-7
Nurcahyo, H., Sumiwi, S. A., Halimah, E., & Wilar, G. (2022). Secondary metabolitm determination from Brebes shallot’s ethanol extract and its ethyl acetate fraction “Allium ascalonicum L.”. Journal of Advanced Pharmacy Education and Research, 12(1), 70-73. doi:10.51847/NfNMFJB9ac
Padervand, M., Lichtfouse, E., Robert, D., & Wang, C. (2020). Removal of microplastics from the environment. A review. Environmental Chemistry Letters, 18(3), 807-828. doi:10.1007/s10311-020-00983-1
Pérez-Nava, J., Hernández-Aldana, F., Martínez-Valenzuela, C., & Rivera, A. (2021). Pseudomonas sp Isolated from Wastewater and their Interaction with Microalgae. Journal of Biochemical Technology, 12(2), 1-5. doi:10.51847/yaQC7KnNoY
Raberg, M., Volodina, E., Lin, K., & Steinbüchel, A. (2018). Ralstonia eutropha H16 in progress: applications beside PHAs and establishment as production platform by advanced genetic tools. Critical Reviews in Biotechnology, 38(4), 494-510. doi:10.1080/07388551.2017.1369933
Ranganadhareddy, A., & Chandrasekhar, Ch. (2021). Production of polyhydroxybutyrate from marine source- A Review. Indian Journal of Ecology,48(6), 1829-1836.
Ravi Teja, K. S., Abraham Peele, K., Venkateswarulu, T. C., Vijetha, P., Krupanidhi, S., & Ranganadha, R. A. (2020). Computational Modeling of Chitinase (Cs CHIO) Protein from Chitiniphilus shinanonensis. Indian Journal of Ecology, 47(11), 151-153.
Raza, Z. A., Abid, S., & Banat, I. M. (2018). Polyhydroxyalkanoates: Characteristics, production, recent developments and applications. International Biodeterioration & Biodegradation, 126, 45-56. doi:10.1016/j.ibiod.2017.10.001
Reddy, A. R., Peele, K. A., Krupanidhi, S., Prabhakar, K. V., & Venkateswarulu,T.C.(2019). Production of Polyhydroxtbutyratye from Acinetobacter nosocomialis RR20 strain using modified mineral salt medium: a statistical approach. International Journal of Environment Science and Technology, 16, 6447–6452.
Requena, R., Vargas, M., & Chiralt, A. (2020). 3 Plastics from bacteria (polyhydroxyalkanoates). Sustainability of Polymeric Materials, edited by Valentina Marturano, Veronica Ambrogi and Pierfrancesco Cerruti, Berlin, Boston: De Gruyter. 39-72.
Ryan, M. P., & Walsh, G. (2016). The biotechnological potential of whey. Reviews in Environmental Science and Bio/Technology, 15(3), 479-498.
Sathya, A. B., Sivasubramanian, V., Santhiagu, A., Sebastian, C., & Sivashankar, R. (2018). Production of polyhydroxyalkanoates from renewable sources using bacteria. Journal of Polymers and the Environment, 26(9), 3995-4012. doi:10.1007/s10924-018-1259-7
Shaheen, R. S., Alsaffan, A. D., Al-Dusari, R. S., Helmi, R. N., & Baseer, M. A. (2022). Self-Perceived Oral Hygiene and Periodontal Health Among Dental and Medical Students, Dentists and Physicians in KSA. Annals of Dental Specialty, 10(1), 126-132. doi:10.51847/NVcZEJ0YBV
Sharma, M., & Dhingra, H. K. (2021). An Overview of Microbial Derived Polyhydroxybutyrate (PHB): Production and Characterization, pp. 143-176. In: Vaishnav A., Choudhary D.K. (eds). Microbial Polymers. Springer, Singapore.
Skariyachan, S., Patil, A. A., Shankar, A., Manjunath, M., Bachappanavar, N., & Kiran, S. (2018). Enhanced polymer degradation of polyethylene and polypropylene by novel thermophilic consortia of Brevibacillus sps. and Aneurinibacillus sp. screened from waste management landfills and sewage treatment plants. Polymer Degradation and Stability, 149, 52-68.
Snoch, W., Stępień, K., Prajsnar, J., Staroń, J., Szaleniec, M., & Guzik, M. (2019). Influence of chemical modifications of polyhydroxyalkanoate-derived fatty acids on their antimicrobial properties. Catalysts, 9(6), 510.
Wu, H., Chen, J., & Chen, G. Q. (2016). Engineering the growth pattern and cell morphology for enhanced PHB production by Escherichia coli. Applied Microbiology and Biotechnology, 100(23), 9907-9916.
Zhang, M., Kurita, S., Orita, I., Nakamura, S., & Fukui, T. (2019). Modification of acetoacetyl-CoA reduction step in Ralstonia eutropha for biosynthesis of poly (3-hydroxybutyrate-co-3-hydroxyhexanoate) from structurally unrelated compounds. Microbial Cell Factories, 18(1), 1-12.
Zhao, F., Gong, T., Liu, X., Fan, X., Huang, R., Ma, T., Wang, S., Gao, W., & Yang, C. (2019). Morphology engineering for enhanced production of medium-chain-length polyhydroxyalkanoates in Pseudomonas mendocina NK-01. Applied Microbiology and Biotechnology, 103(4), 1713-1724. doi:10.1007/s00253-018-9546-8
Zia, K. M., Tabasum, S., Nasif, M., Sultan, N., Aslam, N., Noreen, A., & Zuber, M. (2017). A review on synthesis, properties and applications of natural polymer-based carrageenan blends and composites. International Journal of Biological Macromolecules, 96, 282-301. doi: 10.1016/j.ijbiomac.2016.11.095
Zibi, R. D. N., Tala, V. R. S., Mbopi, P. Y., Bayaga, N. H., Tcheuffa, G. M. N., & Ngoupayo, J. (2022). Comparative Antiplasmodial and Cytotoxic Activities of Coffea arabica and Coffea canephora alkaloids extracts. International Journal of Pharmaceutical and Phytopharmacological Research, 12(1), 54-59. doi:10.51847/md2J5bMnQF