Production of Polyhydroxyalkanoates from Microalgae- A Review
Aluru Ranganadhareddy
Abstract
Polyhydroxyalkanoates are synthesized by microorganisms, similar to synthetic polymers, and completely biodegradable. Polyhydroxyalkanoates or PHAs can be processed in a similar way to synthetic polymers and are considered substitutes for petroleum-based plastics. In the culture media, bacteria and large amounts of organic carbon sources are responsible for 50% of the total cost of production of commercial PHAs. It is only possible for the PHA to have a greater commercial application in a reduction in the cost of production. The potential for microalgae to provide PHAs at a lower cost has been suggested by several studies, they require relatively little nutrients as well as deriving energy from light and CO2, i.e., being photoautotrophs. The purpose of this paper is to examine biopolymer composition and properties, to determine whether petroleum-based plastics can be replaced by PHAs. Microalgal studies that stimulate PHA synthesis are still in the early stages. However, it remains evidentthat microalgae are capable of producing biopolymers at a low cost and can be used to contribute to the environment.
Keywords: Polyhydroxyalkanoate, Microalgae, Bioplastic, Polyhydroxtbutyratye, Biopolymer
Introduction
Over the past seven decades, the production of petroleum-derived plastics has gained considerable attention and is now the most widely used material in industrial and household settings, replacing wood, glass, metals, and building materials (Peng et al., 2020). Lightweight, robust, durable, and resistant to degradation, plastic materials are highly desirable (Saratale et al., 2021). Despite its widespread use and numerous applications, nearly all the plastic in the world is thrown away as garbage or detritus (Emamahadi et al., 2020). There is an annual production of approximately 140 million tons of plastic in the world, requiring about 150 million tons of fossil fuels to be processed and generating large quantities of waste whose decomposition takes thousands of years (Onen Cinar et al., 2020). Since microbial degradation of plastics is extremely difficult, they have proven to be serious environmental problems. As plastics disperse over great distances and end up in sediments, they can remain in the earth’s crust for centuries (Abalansa et al., 2020). The consumption of plastic is expected to increase until 2030 in the coming decades (Cruz et al., 2022). The methods for disposing of plastic materials are challenging (Anjum et al., 2016). Degradation rates are extremely low in landfills. Toxic by-products are generated by incineration. Although recycling is possible, it requires a lot of time and plastic materials might change their properties (Li & Wilkins, 2020). Furthermore, the use of recycled materials is limited by additives like paints and coatings. Because they are biodegradable and do not use fossil fuels, plastics made from synthetic materials can be replaced by biopolymers. At present, however, biodegradable alternatives are still more expensive than petroleum-based polymers, making it a challenge to develop environmentally friendly alternatives for commercial and retail markets (D'Adamo et al., 2021). In addition to their similar material properties to polypropylene, polyhydroxyalkanoates are also completely biodegradable and can be extruded and injected in the same manner (Choi et al., 2020). Composting waste materials derived from them is easy since they degrade relatively fast by soil organisms (Hatti-Kaul et al., 2020). Fermenters are used to produce PHA. Heterotrophic bacteria, including Cupriavidus necator and recombinant Escherichia coli, are responsible for this process (Costa et al., 2018; Tan et al., 2021). In cultivation processes involving organic sources, such as glucose and mineral salts, approximately half of the total production cost is accounted for by organic carbon sources (Price et al., 2020). The cost of manufacturing PHA as a polymer is higher than that of some widely used petroleum-based plastics, limiting its industrial applications (Balaji et al., 2013). Another way to produce PHA is by using the cellular biomass of cyanobacteria or prokaryotic microalgae (Roja et al., 2019). Phytoplankton produces biomass only by converting light and atmospheric CO2 into biomass. Obtaining PHAs at a lower cost can be achieved with a type of microorganism called microalgae. The fact that they are photoautotrophic and have minimal nutrient needs lead to them accumulating PHAs through oxygenated photosynthesis. To increase PHA's market competitiveness and processability, improved polymer characteristics are essential, apart from reducing production costs. Hence, verifying the possible industrial applications of the obtained biopolymers is vital, to learn more about their properties (Costa et al., 2018). There have been several studies that evaluated the properties of biopolymers obtained from microalgae in comparison to synthetic polymers, but very few have addressed molecular mechanisms underlying the process of microalgae synthesizing biopolymers (Asiwe et al., 2022; Dang et al., 2022). Taking a general overview of the production methods of PHAs from microalgae, this review will focus on the properties and compositions of biopolymers, to compare them to petroleum-based plastics.
Microbial Biopolymers
Microbial biopolymers are natural polymers produced and catabolized through numerous organisms which might be non-poisonous to the host and feature a few benefits over petroleum-primarily based plastics (Saratale et al., 2021; Seal et al., 2022). Due to their capacity for utility and brief decomposition through microorganisms inclusive of bacteria, biopolymers are revolutionary and exciting. In microbial cells below stress, those biopolymers collect as reserve substances (Onen et al., 2020). Biologically derived PHAs have proven enormous potential in several fields inclusive of pharmaceuticals, agriculture, biofuel, medicine, disposable, and chromatography (Chen & Zhang, 2018). In many microorganisms, polyhydroxyalkanoates (PHA) polyesters are produced and accumulated, commonly once they enter the desk-bound section of improvement (Bhola et al., 2021). PHAs consist of intracellular inclusions which could keep carbon, energy, and account for as much as 80% of a mobileular's weight (Roja et al., 2019; Chakraverty & Bondyopadhyay, 2022). When different important factors inclusive of oxygen, phosphorus, or nitrogen are scarce, they're produced inside the mobileular as insoluble cytoplasmic inclusions inside the presence of excessive carbon. Because those polymeric substances do now no longer considerably alter the mobileular's osmotic state, they may be saved at excessive concentrations in the mobileular (Zhang et al., 2018; AlHussain et al., 2022). These inclusions are spherical, with suggest sizes of 0.2-0.7 m, and encased in a 2 nm membrane of round 2% protein and 0.5% lipids. A phospholipid layer, polymerases, depolymerases, and nonspecific cytosolic proteins all play a function inside the granules (Hamad et al., 2018). The lipid monolayer is important to avoid changing the amorphous to the crystalline polyester state, which might be a lot stable but could induce cell rupture (Galiano et al., 2018; Burlaka, 2022; Govindaraj et al., 2022). In their work on bacterial polyhydroxyalkanoate granules, Grage et al. confirmed the presence of a phospholipid layer in PHA isolates (Grage et al., 2009). The authors assume that the granules have a polyester core by a physical phenomenon with integrated or connected proteins appreciate the PHA synthase, phasins, depolymerizing enzymes, and restrictive proteins additionally to the monolayer of phospholipids. Notwithstanding the growing facts, the presence of a phospholipid coat in vivo has nevertheless been proven. Various findings have forged doubt on the presence of the supermolecule layer in vivo, notably from the microscopy investigations, which recommend that the lipid coat could type as a result of associate in nursing experimental error throughout PHA extraction and processing (Obruca et al., 2020; Remizova et al., 2022).
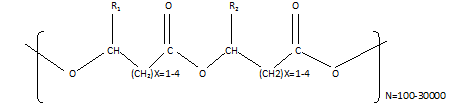
Thermoplastic polymers composed of linear hydroxyalkanoic acid are called PHAs. As one monomeric unit's carboxyl group bonds to another monomeric unit's hydroxyl group, an ester bond is formed (Ranganadha & Chandrasekhar, 2021). Figure 1 shows how they are structured with "n" reaching 35,000 monomers. Monomers of PHA have various R groups from hydrogen atoms to methyl tridecyl (Reddy et al., 2019). The alkyl side chains of phosphoric acid contain usually a saturated alkyl group. However, they can also show a variety of chemical structures that include alkylated, unsaturated, aromatic, branched, epoxidized, and substituted alkyl groups. PHA thermoplastics' side chains can be chemically modified by cross-linking unsaturated bonding (Reddy et al., 2017).
|
|
|
Figure 1. Structure of Polyhydroxyalkanoates with R1 and R2 are alkyl groups (C-C) |
Essentially, both short-chain and medium-chain hydroxyalkanoic acids have carbon chains of around three to five carbon atoms; medium-chain acids have carbon chains of about six to fourteen, and long-chain acids have carbon chains of more than 14 atoms. PHAs are divided into three groups based on the number of carbon atoms in their carbon chain: short-chain hydroxyalkanoic acids (PHASCL), medium-chain hydroxyalkanoic acids (PHAMCL), and long-chain hydroxyalkanoic acids (PHALCL) (Saratale et al., 2021). Among these groups, PHA synthases differ primarily due to their substrate specificity, which allows them to accept three hydroxy acids with a specific carbon length range. Polymers and copolymers can thus be made from different substrates and metabolites depending on the microorganism. PHAMCL is elastomeric and rubber-like, in contrast to PHASCL, which is comparable to traditional plastics (Ranganadha & Chandrasekhar, 2021). A commonly used commercial form of PHA is PHB (poly-3-hydroxybutyrate). Bacterial bioplastics are generally more expensive than petroleum-based polymers such as polyethene, polypropylene, and high-density polyethylene which is a limiting factor for their use in the industrial sector. PHA manufacturing costs are often increased due to the high cost of carbon sources (Reddy et al., 2019; AlShehri et al., 2022; Mohandas et al., 2022). More than one hundred microalgae strains are examined for the generation of PHAs. Microalgae use sunshine, carbon dioxide, water, and nutrients in this chemical process cycle to form PHAs collected, purified, and processed in an exceedingly big selection of plastic products. PHAs can be deployed in active microorganism habitats once usage and therefore the microbial degradation of such a compound produce carbonic acid gas and water that microalgae devour (Van-Thuoc et al., 2008).
Properties and Composition of PHA’s
Thanks to the chemical heterogeneousness of their radicals, PHAs exhibit properties that are just like those of ancient polymers (Maghami et al., 2022; Ranganadhareddy, 2022). These polymers will vary from onerous and brittle thermoplastics to elastomers, rubbers and adhesives supported the chemical compound makeup (Albuquerque & Malafaia, 2018). They possess a degree of polymerization of up to 30,000 supported by their huge molecular masses. The snap of a polymer rises because the chain length or the amount of comonomers in it grows; therefore, PHAs have varied characteristics looking on their monomeric composition. The molecular mass, thermal characteristics, and crystallinity index are the foremost researched physical parameters of PHAssince they confirm the polymer's processability as shown in Table 1. A polymer molecular mass and relative molecular mass distribution are crucial qualities for industrial applicability and polymers with molecular lots not up to 4X104Da have poor mechanical properties (Ertan et al., 2021). The molecular mass of those molecules ranges between 2X105 to 3X105 Da, supported the microorganism species utilized and growth circumstances reminiscent of pH, conditions, culturing modes, and therefore the kind and concentration of the carbon source. The extraction procedure wedged the molecular mass of PHAs as well (Reddy et al., 2019). Laycock et al. planned that the molecular mass proportion of biopolymers is connected to their terminal qualities via molecule structural regulations (Laycock et al., 2013). The size of the chemical compound chains determines the PHA characteristics, whereas structural rearrangements are influenced by the degree of polymerization. At room temperature, a polymer’s thermal characteristics (Tm – melting temperature, Tg – glass transition temperature) and qualities like crystallinity and crystallization time determine specific mechanical characteristics and are also crucial markers for thermal processing. Depending on the polymer composition, PHAs have a melting temperature ranging from 50 to 180 degrees Celsius and 30 to 70% crystallinity. PHAs possessing crystallinity around 60 and 80 percent, according to Assis et al., are found to be rigid. Accordingly, the polymer lengths of PHAs that are malleable and highly pliable are moderate (30 to 40%) and short (30%). PHA’s processing qualities are improved by its reduced degree of crystallinity, enhancing the range of viable industrial uses (Thulasidharan et al., 2021).
Table 1. Physical and thermal properties of different types of PHAs (Costa et al., 2019)
|
Properties |
PHB |
PHBV |
PP |
HDPE |
|
Crystallinity (%) |
60-80 |
30-40 |
68 |
60-80 |
|
Melting temperature (oC) |
37 |
45 |
174 |
125-132 |
|
Molecular mass (KDa) |
180 |
164 |
190 |
200-600 |
|
Breaking strength (%) |
4.9 |
72-87 |
400 |
12 |
The monomeric composition of PHAs is highly reliant on growing conditions and the originating strain, and it is specifically connected to the polymer’s characteristics and potential commercial uses (Roja et al., 2019). Despite the bacteria’s ability to acquire more PHAs, the polymers produced are dominated by 4-10 carbon atom monomers (Costa et al., 2018). Large proportions of these monomers give PHAs characteristics that make them unsuitable for industrial use. PHAs have high applicability in various fields such as agriculture, biomedical sciences, and environment, apart from their usual usage in applications such as bag making, fabrics, and so on. With PHA, disposable products, artificial bones, blood vessel substitutes, and heart valve tissue engineering materials can all be made (Thulasidharan et al., 2021).
PHAs Production in Microalgae and Biosynthetic Pathways
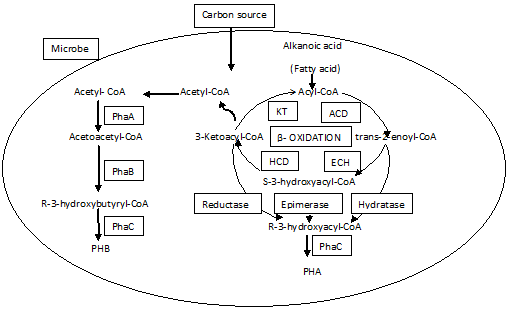
According to numerous studies, PHA synthesis by microalgae is known to occur when microbes grow in a nutrient-deficient condition. When microalgae thrive in surroundings deficient in nitrogen and/or phosphorus, their metabolic pathways are altered to synthesize carbon-rich molecules like PHAs, which operate as a source of energy storage (Mendhulkar & Shetye. 2017). Costa et al. (2018) found that the cyanobacteria Synechococcus subsalsus and Spirulina sp. LEB-18 are exposed to a nitrogen deficient state; they shift carbon to other metabolic pathways and build biopolymers that can be employed as carbon and energy storage molecules when conditions improve. Various cultivation tactics are used to enhance the biopolymer proportion of PHAs, including the usage of reduced or increased media of specific nutrients, including phosphate, nitrogen, or concurrent deprivation of phosphorus and nitrogen, the inclusion of acetate and propionate, along with higher salinity, restriction of gas exchange, and waste-water usage as sewage, among others. The most frequent metabolic route, which may be seen in a wide spectrum of bacteria that possess an ability to synthesize PHA and fatty acids, is linked to acetyl CoA, a prevalent precursor. The generation of PHAs by microalgae is thought to follow a similar metabolic mechanism. The PHA is derived from acetyl-coenzyme A (acetyl-CoA) by three enzymatic processes as shown in Figure 2 (Li et al., 2021; Ranganadhareddy, 2022).
|
|
|
Figure 2. Biosynthetic pathway of Polyhydroxtbutyratye in microbes |
3- Ketothiolasecatalyzes the conversion of two acetyl-CoA molecules as one acetoacetyl-CoA molecule, NADPH-dependent acetoacetyl-CoA reductase catalyzes the reduction of acetoacetyl-CoA to D-3-hydroxybutyryl-CoA and PHA synthase facilitates the association of the D-3-hydroxybutyryl and a PHA molecule through an ester linkage. 3-hydroxy fatty acids are found in β, as well as numerous other PHAs. Between one methyl to thirteen tridecyl carbons make up the pendant group. There are fatty acids having hydroxyl groups at positions 4, 5, and 6 as well as pendent groups carrying substituents or unsaturation, which result in various PHA copolymers and homopolymers (Ranganadha & Chandrasekhar, 2021; Knobloch et al., 2022).
Large scale Production of PHAs from Microalgae
Spirulina sp. was discovered in 1519 in Spain and the study of microalgae and their health benefits increased as a result of the observation of their consumption by Aztecs in the Valley of Mexico, the fact that flamingos survive by eating algae, and flamingos consuming algae as a result of scientific evidence (Sudhakar et al., 2018). An initial processing plant was established in 1969 to commercialize Spirulina sp., primarily for direct consumption (Cardias et al., 2018). In contrast to the production of microalgae used directly for food, bioplastics produced from microalgae PHAs are not yet commercially available, even though numerous firms are involved in this process, depending on the country, at various stages (Tabet et al., 2018). A typical microalgae provides valuable compositions such as polysaccharides, lipids, proteins, antioxidants, vitamins, pigments, minerals, fatty acids, polyhydroxyalkanoate, micronutrients, etc. A large number of high-value products can be synthesized from the microalgae biomass, including PHAs, which can be used in exceptional commercial applications, microalgae biomass also contains different compounds of high added value, making it possible to commercially acquire highly valuable products (Cardias et al., 2018). According to Singh and Mallick, these demanding situations had to be addressed for making cyanobacterial PHAs a business reality: harvesting of cyanobacterial biomass, drying cyanobacterial biomass, and cyanobacterial mass cultivation. Despite their potential to replace 33% of polymeric materials in the market, PHAs still have a limited market. In upcoming decades, technological improvements in many components in combination with improvements in biological and artificial biology systems will be required to create cyanobacterial species capable of much greater photosynthetic and PHA production (Singh & Mallick, 2017). PHA studies are not only about finding the appropriate biomass and converting it into bioplastic, but also about making it economically sustainable on an industrial scale. The cyanobacteria, in addition to their value as a source of novel and biochemically active herbal products, are additionally being identified as one of the most promising candidates (Kardile & Shirsat, 2020). In the coming years, microalgae production is predicted to keep growing in the global market, resulting in the expansion of the scale of this industry. It is anticipated that biorefineries will convert all biomass compounds into high-value new products identical to those produced by petrochemical refineries.
Conclusion
Sourcing PHAs from microalgae can be viewed as an essential method for lowering polymer acquisition costs and as a result, increasing competitiveness versus synthetic polymers. Microalgae are a viable source for PHAs since these are the only microbes that use photosynthesis to collect PHAs, employing CO2 and light as their primary sources of energy along with having the low nutritional need for their growth. Microbes consume CO2 to reduce the warming impact caused by industrial CO2 emissions. These microbes play an essential role in protecting the environment. In essence, microalgae synthesis of PHAs minimizes the consumption of fossil fuel resources, as well as reducesCO2 emissions, thereby reducing the environmental impact of the process. Despite, still not knowing how PHAs are produced in microalgae, the growth conditions, species, and extraction methods of these polymers all affect their characteristics, which are crucial for their commercialization. Microalgae-stimulated PHA synthesis is currently in the early stages of research, and the PHAs produced are not yet commercially available. Regardless, microalgae can create biopolymers at a cheaper cost and play an essential role in the environment.
Acknowledgments: None
Conflict of interest: None
Financial support: None
Ethics statement: None
Abalansa, S., El Mahrad, B., Vondolia, G. K., Icely, J., & Newton, A. (2020). The marine plastic litter issue: a social-economic analysis. Sustainability, 12(20), 8677. doi:10.3390/su12208677
Albuquerque, P. B., & Malafaia, C. B. (2018). Perspectives on the production, structural characteristics and potential applications of bioplastics derived from polyhydroxyalkanoates. International Journal of Biological Macromolecules, 107, 615-625. doi:10.1016/j.ijbiomac.2017.09.026.
AlHussain, B. S., AlFantoukh, M. A. M., Alasmari, K. M. A., AlHrab, F. A., Alotaibi, F. A., Alaybani, W. H., & AlOtaibi, I. A. B. (2022). Clinical knowledge of orthodontics complication and emergencies among interns and dentists in Riyadh City. Annals of Dental Specialty, 10(2), 45-51. doi:10.51847/9H6hhUJo8E
AlShehri, O. M., Jali, N. M., Almutairi, Y. M., Aljrais, M. M., Alsirhani, A. M., & AlQudairi, A. S. (2022). Common causes of cusp fracture in adults; a systematic review. Annals of Dental Specialty, 10(2), 35-38. doi:10.51847/jBTdUqt7Cx
Anjum, A., Zuber, M., Zia, K. M., Noreen, A., Anjum, M. N., & Tabasum, S. (2016). Microbial production of polyhydroxyalkanoates (PHAs) and its copolymers: a review of recent advancements. International Journal of Biological Macromolecules, 89, 161-174. doi:10.1016/j.ijbiomac.2016.04.069
Asiwe, N., Asiwe, J. N., Asiwe, T. N., & Asiwe, P. C. (2022). Awareness of COVID-19 and its vaccine acceptability among young adult population of agbor, Delta State, Nigeria. International Journal of Pharmaceutical and Phytopharmacological Research, 12(2), 24-29. doi:10.51847/tvmGc5ytyZ
Balaji, S., Gopi, K., & Muthuvelan, B. (2013). A review on production of poly β hydroxybutyrates from cyanobacteria for the production of bio plastics. Algal Research, 2(3), 278-285. doi:10.1016/j.algal.2013.03.002
Bhola, S., Arora, K., Kulshrestha, S., Mehariya, S., Bhatia, R. K., Kaur, P., & Kumar, P. (2021). Established and emerging producers of PHA: Redefining the possibility. Applied Biochemistry and Biotechnology, 193(11), 3812-3854. doi:10.1007/s12010-021-03626-5
Burlaka, I. (2022). Approach to correction of apoptotic disorders in children with early diabetic nephropathy. Journal of Advanced Pharmacy Education and Research, 12(2), 104-109. doi:10.51847/G6i3231Jnm
Cardias, B. B., de Morais, M. G., & Costa, J. A. V. (2018). CO2 conversion by the integration of biological and chemical methods: Spirulina sp. LEB 18 cultivation with diethanolamine and potassium carbonate addition. Bioresource Technology, 267, 77-83. doi:10.1016/j.biortech.2018.07.031
Chakraverty, R., & Bondyopadhyay, J. (2022). Recent insights into the association between stress, anxiety and hypertension in adults: a systematic review. International Journal of Pharmaceutical and Phytopharmacological Research, 12(2), 12-17. doi:10.51847/Bug18QDArj
Chen, G. Q., & Zhang, J. (2018). Microbial polyhydroxyalkanoates as medical implant biomaterials. Artificial Cells, Nanomedicine, and Biotechnology, 46(1), 1-18. doi:10.1080/21691401.2017.1371185
Choi, S.Y., Rhie, M.N., Kim, H.T., Joo, J.C., Cho, I.J., Son, J., Jo, S.Y., Sohn, Y.J., Baritugo, K. A., Pyo, J., et al. (2020). Metabolic engineering for the synthesis of polyesters: a 100-year journey from polyhydroxyalkanoates to non-natural microbial polyesters. Metabolic Engineering, 58, 47-81. doi:10.1016/j.ymben.2019.05.009
Costa, S. S., Miranda, A. L., de Morais, M. G., Costa, J. A. V., & Druzian, J. I. (2019). Microalgae as source of polyhydroxyalkanoates (PHAs)—A review. International Journal of Biological Macromolecules, 131, 536-547. doi:10.1016/j.ijbiomac.2019.03.099.
Costa, S.S., Miranda, A.L., Andrade, B.B., de Jesus Assis, D., Souza, C.O., de Morais, M.G., Costa, J.A. V., & Druzian, J.I. (2018). Influence of nitrogen on growth, biomass composition, production, and properties of polyhydroxyalkanoates (PHAs) by microalgae. International Journal of Biological Macromolecules, 116, 552-562. doi:10.1016/j.ijbiomac.2018.05.064
Cruz, R. A., Oehmen, A., & Reis, M. A. (2022). The impact of biomass withdrawal strategy on the biomass selection and polyhydroxyalkanoates accumulation of mixed microbial cultures. New Biotechnology, 66, 8-15. doi:10.1016/j.nbt.2021.08.004
D'Adamo, S., Kormelink, R., Martens, D., Barbosa, M. J., & Wijffels, R. H. (2021). Prospects for viruses infecting eukaryotic microalgae in biotechnology. Biotechnology Advances, 54, 107790. doi:10.1016/j.biotechadv.2021.107790
Dang, B.T., Bui, X.T., Tran, D.P., Ngo, H.H., Nghiem, L.D., Nguyen, P.T., Nguyen, H.H., Lin, C., Lin, K.Y. A., &Varjani, S. (2022). Current application of algae derivatives for bioplastic production: A review. Bioresource Technology, 347, 126698. doi:10.1016/j.biortech.2022.126698
Emamhadi, M. A., Sarafraz, M., Akbari, M., Fakhri, Y., Linh, N. T. T., & Khaneghah, A. M. (2020). Nanomaterials for food packaging applications: a systematic review. Food and Chemical Toxicology, 146, 111825. doi:10.1016/j.fct.2020.111825.
Ertan, F., Keskinler, B., & Tanriseven, A. (2021). Exploration of Cupriavidus necator ATCC 25207 for the Production of Poly (3-hydroxybutyrate) Using Acid Treated Beet Molasses. Journal of Polymers and the Environment, 29(7), 2111-2125. doi:10.1007/s10924-020-02020-2
Galiano, F., Briceño, K., Marino, T., Molino, A., Christensen, K. V., & Figoli, A. (2018). Advances in biopolymer-based membrane preparation and applications. Journal of Membrane Science, 564, 562-586. doi:10.1016/j.memsci.2018.07.059
Govindaraj, A., Paulpandian, S. S., & Shanmugam, R. (2022). A comparative assessment of the impact of citrullus lanatus rind and pulp extracts on Streptococcus mutans. International Journal of Dental Research and Allied Sciences, 2(2), 1-6. doi:10.51847/ffD4G8LgXK
Grage, K., Jahns, A. C., Parlane, N., Palanisamy, R., Rasiah, I. A., Atwood, J. A., & Rehm, B. H. (2009). Bacterial polyhydroxyalkanoate granules: biogenesis, structure, and potential use as nano-/micro-beads in biotechnological and biomedical applications. Biomacromolecules, 10(4), 660-669. doi:10.1021/bm801394s
Hamad, K., Kaseem, M., Ayyoob, M., Joo, J., & Deri, F. (2018). Polylactic acid blends: The future of green, light and tough. Progress in Polymer Science, 85, 83-127. doi:10.1016/j.progpolymsci.2018.07.001
Hatti-Kaul, R., Nilsson, L. J., Zhang, B., Rehnberg, N., & Lundmark, S. (2020). Designing biobased recyclable polymers for plastics. Trends in Biotechnology, 38(1), 50-67. doi:10.1016/j.tibtech.2019.04.011
Kardile, D., & Shirsat, M. (2020). Synthesis and in vitro evaluation of coupled mercaptobenzimidazole derivatives used as a potent biological agent. International Journal of Pharmaceutical and Phytopharmacological Research, 10(1), 127-133.
Knobloch, J., Müller, C., & Hildebrandt, J. P. (2022). Expression levels and activities of energy-yielding ATPases in the oligohaline neritid snail Theodoxus fluviatilis under changing environmental salinities. Biology Open, 11(2), bio059190. doi:10.1242/bio.059190
Laycock, B., Halley, P., Pratt, S., Werker, A., & Lant, P. (2013). The chemomechanical properties of microbial polyhydroxyalkanoates. Progress in Polymer Science, 38(3-4), 536-583. doi:10.1016/j.progpolymsci.2012.06.003
Li, M., & Wilkins, M. R. (2020). Recent advances in polyhydroxyalkanoate production: Feedstocks, strains and process developments. International Journal of Biological Macromolecules, 156, 691-703. doi:10.1016/j.ijbiomac.2020.04.082
Li, M., Li, K. & Ren, Y. (2021). Nesfatin-1 protects H9c2 cardiomyocytes against cobalt chloride-induced hypoxic injury by modulating the MAPK and Notch1 signaling pathways. Journal of Biological Research-Thessaloniki, 28(21). doi:10.1186/s40709-021-00147-4
Maghami, H., Parhizkar, H., Riasaty, A., Banani, A., Poustfroosh, M., & Hasehmpour-Sadeghian, M. (2022). Identifying the parenting style of students’ parents of an school at Shiraz University of medical sciences. Journal of Advanced Pharmacy Education and Research, 12(1), 91-96. doi:10.51847/tLcoO2yvKI
Mendhulkar, V. D., & Shetye, L. A. (2017). Synthesis of biodegradable polymer polyhydroxyalkanoate (PHA) in cyanobacteria Synechococcus elongates under mixotrophic nitrogen-and phosphate-mediated stress conditions. Industrial Biotechnology, 13(2), 85-93. doi:10.1089/ind.2016.0021
Mohandas, R., Ramani, P., & Mohapatra, S. (2022). Exploring Corono-condylar distance as a radiographic marker for chronological age. International Journal of Dental Research and Allied Sciences, 2(2), 7-9. doi:10.51847/xF069fnRvk
Obruca, S., Sedlacek, P., Slaninova, E., Fritz, I., Daffert, C., Meixner, K., Sedrlova, Z., & Koller, M. (2020). Novel unexpected functions of PHA granules. Applied Microbiology and Biotechnology, 104(11), 4795-4810. doi:10.1007/s00253-020-10568-1
Onen Cinar, S., Chong, Z. K., Kucuker, M. A., Wieczorek, N., Cengiz, U., & Kuchta, K. (2020). Bioplastic production from microalgae: a review. International Journal of Environmental Research and Public Health, 17(11), 3842. doi:10.3390/ijerph17113842
Peng, B.Y., Li, Y., Fan, R., Chen, Z., Chen, J., Brandon, A.M., Criddle, C. S., Zhang, Y., & Wu, W.M. (2020). Biodegradation of low-density polyethylene and polystyrene in superworms, larvae of Zophobas atratus (Coleoptera: Tenebrionidae): Broad and limited extent depolymerization. Environmental Pollution, 266, 115206. doi:10.1016/j.envpol.2020.115206.
Price, S., Kuzhiumparambil, U., Pernice, M., & Ralph, P. J. (2020). Cyanobacterial polyhydroxybutyrate for sustainable bioplastic production: critical review and perspectives. Journal of Environmental Chemical Engineering, 8(4), 2213-3437. doi:10.1016/j.jece.2020.104007
Ranganadhareddy, A. (2022). A Review on Production of Polyhydroxyalkanoates in Microorganisms. Journal of Biochemical Technology, 13(1), 1-6. doi:10.51847/Uo3EEbmgID
Ranganadhareddy, A., & Chandrasekhar, Ch. (2021). Production of polyhydroxybutyrate from marine source- A Review. Indian Journal of Ecology, 48(6), 1829-1836.
Reddy, A. R., Kumar, R. B., & Prabhakar, K. V. (2017). Isolation and Identification of PolyHydroxyButyrate (PHB) producing bacteria from Sewage sample. Research Journal of Pharmacy and Technology, 10(4), 1065-1069. doi:10.5958/0974-360X.2017.00193.7
Reddy, A. R., Peele, K. A., Krupanidhi, S., Prabhakar, K. V., & Venkateswarulu, T. C. (2019). Production of polyhydroxybutyrate from Acinetobacter nosocomialis RR20 strain using modified mineral salt medium: a statistical approach. International Journal of Environment Science and Technology, 16, 6447–6452. doi:10.1007/s13762-018-2102-3
Remizova, A. A., Bitarov, P. A., Epkhiev, A. A., & Remizov, N. O. (2022). Reparative-regenerative features of bone tissue in experimental animals treated with titanium implants. Journal of Advanced Pharmacy Education and Research, 12(2), 110-116. doi:10.51847/Sprxb1DKyv
Roja, K., Sudhakar, D. R., Anto, S., & Mathimani, T. (2019). Extraction and characterization of polyhydroxyalkanoates from marine green alga and cyanobacteria. Biocatalysis and Agricultural Biotechnology, 22, 101358. doi:10.1016/j.bcab.2019.101358
Saratale, R. G., Cho, S. K., Saratale, G. D., Kadam, A. A., Ghodake, G. S., Kumar, M., Bharagava, R. N., Kumar, G., Kim, D. S., Mulla, S. I., et al. (2021). A comprehensive overview and recent advances on polyhydroxyalkanoates (PHA) production using various organic waste streams. Bioresource Technology, 325, 124685. doi:10.1016/j.biortech.2021.124685
Seal, T., Chaudhuri, K., & Pillai, B. (2022). Wild edible plants of Meghalaya state in India: assessment of nutritional and toxicological potential. International Journal of Pharmaceutical and Phytopharmacological Research, 12(2), 1-11. doi:10.51847/kUHnRyZSOe
Singh, A. K., & Mallick, N. (2017). Advances in cyanobacterial polyhydroxyalkanoates production. FEMS Microbiology Letters, 364(20), 1-13. doi:10.1093/femsle/fnx189
Sudhakar, K., Mamat, R., Samykano, M., Azmi, W. H., Ishak, W. F. W., & Yusaf, T. (2018). An overview of marine macroalgae as bioresource. Renewable and Sustainable Energy Reviews, 91, 165-179. doi:10.1016/j.rser.2018.03.100
Tabet, A., Boukhari, A., & Nouidjem, Y. (2018). Phenolic content, HPLC analysis and antioxidant activity extract from Tamarix articulata. Journal of Advanced Pharmacy Education & Research, 8(4), 1-8.
Tan, D., Wang, Y., Tong, Y., & Chen, G. Q. (2021). Grand challenges for industrializing polyhydroxyalkanoates (PHAs). Trends in Biotechnology, 39(9), 953-963. doi:10.1016/j.tibtech.2020.11.010
Thulasidharan, D., Arumugam, A., & Uppuluri, K. B. (2021). Research and economic perspectives on an integrated biorefinery approach for the simultaneous production of polyhydroxyalkanoates and biohydrogen. International Journal of Biological Macromolecules, 193, 1937-1951. doi:10.1016/j.ijbiomac.2021.11.025
Van‐Thuoc, D., Quillaguaman, J., Mamo, G., & Mattiasson, B. (2008). Utilization of agricultural residues for poly (3‐hydroxybutyrate) production by Halomonas boliviensis LC1. Journal of Applied Microbiology, 104(2), 420-428. doi:10.1111/j.1365-2672.2007.03553.x
Zhang, J., Shishatskaya, E. I., Volova, T. G., da Silva, L. F., & Chen, G. Q. (2018). Polyhydroxyalkanoates (PHA) for therapeutic applications. Materials Science and Engineering: C, 86, 144-150. doi:10.1016/j.msec.2017.12.035