Effect of Lysine and Histidine Residues Modification on the Voltage Gating of the Mitochondrial Porin
Jalal Aljamal
Abstract
Mitochondrial porin is a complex channel with many diverse functional properties. Therefore, along with measuring single-channel conductance and selectivity, monitoring VDAC's voltage gating has become the gold standard for assessing mitochondrial porin function. To understand the mechanisms of gating and ion selectivity, the detergent-solubilized porin was incubated with different concentrations of diethylpyrocarbonate (DEPC). The extent of the reaction of DEPC with histidine residues was monitored by differential absorbance at 242 nm. The resulting modified proteins were loaded onto (CM-cellulose) columns, and the collected fractions were examined for their activity. The apparent ion selectivity was determined by measuring the membrane potential at zero-current conditions. The reversal potentials obtained for the unmodified-porin and ethoxyformyl-porin eluted from the resin (CMC-52) with the equilibration buffer were, respectively, +7 mV and -6 mV. The activity was slowly restored by incubation with hydroxylamine, and the extent of reversal depended on the reagent's concentration. The histidine residues are non-essential in the voltage-gating process since their modification has no direct impact on porin inactivation. However, the decrease of both the channel conductance and the critical potential needed to close the channel resulted from the covalent modification of one or more lysine residues in the EF-porin fraction, suggesting that lysine residues may contribute, in a unique way, to the gating process.
Keywords: VDAC, Chemical modifications, Voltage-gating, Mitochondria, Diethylpyrocarbonate
Introduction
The voltage-dependent anion-selective channel (VDAC) forms the central diffusion pore of the mitochondrial outer membrane. VDAC1, the main isoform in mitochondria, is the most abundant and one of the most extensively studied proteins of the mitochondrial outer membrane. VDAC1 as outer-inner membrane coupling factor, allows the exchange of small molecules up to 5 kDa, including nucleotides, and other metabolites across the mitochondrial outer membrane (Ellenrieder et al., 2019; Endo & Sakaue, 2019). However, since its discovery in the mid-1970s, elucidating the mechanisms underlying voltage-gating events remains challenging. The N-terminal domain is thought to help stabilize the pore's structure by its interaction with the channel's wall (Reif et al., 2019; Najbauer et al., 2022), but at the same time, it is considered the mobile part of the protein, being exposed to the cytosol under certain conditions (Zachariae et al., 2012; Bohm et al., 2020). In general, VDAC mainly assumes a high conductance or open state in the absence of electrical potential, with a single channel conductance of about 3.7 nS in 1 M KCl.
On the other hand, VDAC switches to low conductance or closed states at high positive or negative voltages, with transmembrane potentials mid-points typically around ±30 mV. These states are characterized by a conductance of roughly 2 nS vs. 4 nS in 1M KCl and a slight change in ion selectivity (Rappaport et al., 2015; Noskov et al., 2016). Dynamic simulations indicate that these transitions are controlled by changes in the disordered N-terminal of the pore-forming protein (Preto & Krimm, 2021). Thus, VDAC's voltage dependence stems from the fact that as the membrane potential increases to move the excess gating charge of the voltage sensor, the probability of channel opening will decrease.
According to reconstitution experiments with planar lipid bilayer membranes, the mitochondrial outer membrane pore has an estimated effective diameter of approximately 2–2.8 nm in the open state (Colombini, 2016), which agrees well with the electron microscopic analysis of mitochondrial outer membranes and NMR and X-ray structures (Jaremko et al., 2016; Zeth & Zachariae, 2018; Najbauer et al., 2021) and cryoelectron microscopy (cryoEM) blotting methods (Martynowycz et al., 2020). There are already more than 100 sequences of eukaryotic porins identified. (Kayser et al., 1989; Benz, 2021). The channel properties, such as single-channel conductance, voltage dependence, and selectivity, are essentially the same. However, sequence comparison of porins shows no apparent homology. Nonetheless, there is a high homology in their predicted structural folds (Gattin et al., 2015; Höhr et al., 2018; Diederichs et al., 2021).
VDAC is also somewhat selective for anions over cations of comparable size and charge (Colombini, 2016), suggesting that fixed positively charged side chains are present in or near the pore. Therefore, the prediction of the possible identities of the implicated residues in the gating mechanism associated with the protein channel could be restricted to the side chains of the basic amino acids: histidine, lysine, and arginine.
A series of mutagenesis and chemical modification studies identified several residues throughout the protein sequence that influence gating behavior. This finding led to the development of a gating model in which the VDAC gating process is mediated by an outward movement of a positively charged region called the voltage sensor, resulting in a smaller pore and reversal of the channel selectivity (Thomas et al., 1993; Aljamal, 2002).
The histidine side chain plays a critical role in protein stability. This residue has been shown to stabilize proteins through numerous aromatic and molecular interactions with adjacent residues (Martfeld et al., 2016). Diethylpyrocarbonate (DEPC) has primarily been used to investigate the role of histidine residues in protein functioning because it allows the study of the proteins under native conditions (Biehn et al., 2021). However, it has to be taken into account that DEPC is not selective to histidine residue and modifies other amino acid residues such as Lys and occasionally Arg. Furthermore, most modifications are reversible and can be removed by nucleophilic agents such as hydroxylamine. However, the bis-modification of histidine residues is irreversible (Zhou & Vachet, 2012).
Consequently, the primary goal of the current investigation was to assess the importance of the histidine residues located on the hydrophilic domains of the LDAO-porin in the phenomenon of voltage-gating and anion selectivity. Therefore, experiments on reversible chemical modification were started using diethylpyrocarbonate to look into the potential function of the histidine residues.
Materials and Methods
Hydroxyapatite powder (Bio-gel HTP) was purchased from Bio-Rad Laboratories; lauryl-(dimethyl)-amine oxide (LDAO), acrylamide, and N, N-methylene bisacrylamide from Serva; and molecular mass markers for SDS-PAGE from Boehringer Mannheim. Celite 535 and diethylpyrocarbonate were purchased from Roth and Sigma, respectively. All the other salts and buffers used were of analytical grade and purchased from Merck.
Purification of Bovine Heart Mitochondrial Porin
Mitochondria were isolated from the bovine heart by differential centrifugation. Following the removal of soluble proteins by hypotonic shock and centrifugation, porin protein was purified by solubilizing the mitochondrial membranes with 2% lauryl dimethyl amine oxide (LDAO) and chromatography on a hydroxyapatite/celite (ratio 2:l) column. The porin was eluted with the solubilization buffer containing 2 mM KH2PO4 pH 7.2, 16 mM KCl, and a fraction collector was used. The isolated protein was dialyzed and concentrated to a volume of approximately 5 ml using an Amicon apparatus against a solubilization buffer. Carboxymethyl-cellulose Cation Exchange Chromatography The dialyzed LDAO-porin was loaded onto a carboxymethyl-cellulose (CMC52) cation exchange column equilibrated with an equilibration buffer. The column was washed with 3 ml of equilibration buffer, the first three fractions were discarded, and the retained proteins were eluted by increasing the ionic strength of the eluting buffer with 5 mM KPi pH 7.2, 50 mM KCl. All collected fractions were examined for their activity and protein electrophoresis.
Gel Electrophoresis
Protein purity analysis was carried out using Tris-Glycine SDS-polyacrylamide gel electrophoresis with 16 and 0.4% of acrylamide and bisacrylamide, respectively, as previously described (Laemmli, 1970). Proteins in LDAO-containing fractions were precipitated with cold acetone, incubated in an SDS sample buffer for 5 minutes at 100ºC, and placed on top of 16% polyacrylamide gels for separation. Power was set to ∼100 V constant voltage until the bromophenol blue reached the end of the gel. Gels were stained using 0.25% Coomassie brilliant blue R-250 and subsequent destaining in methanol, and acetic acid solutions.
Porin Histidine Ethoxyformylation
Stock Diethylpyrocarbonate (DEPC) solution (6.71 M) was diluted in absolute ethanol to working concentrations. In addition, LDAO-porin was diluted into 2% LDAO, 1 mM EDTA, and 10 mM Tris-HCl, pH 7.2 (equilibration buffer) to a final concentration of (40– 60 mg porin /ml) prior to each experiment.
LDAO-porin was incubated at 4°C with 0.1–20 mM DEPC. The reaction was initiated by adding DEPC and concluded when the absorbance maximum at 242 nm (ε = 3000 M-1 cm-1) was obtained (Ambasht, 2020). Aliquots of a sample solution containing DEPC were analyzed at various intervals during the incubation. The reaction was then subjected to cation-exchange chromatography using a CMC-52 column. Next, the carbethoxy group was removed from histidyl residues by incubating the EF-porin with 0.33 M hydroxylamine, pH 7.5, at 4°C for 0.5–24 hrs. In a Perkin-Elmer spectrophotometer, the ethoxyformylation and deacylation of histidine residues were monitored spectrophotometrically between 320 and 210 nm (Hunsicker-Wang & Konkle, 2018).
Porin Reconstitution and Electrophysiological Recording
Porin was isolated from bovine heart mitochondrial membranes, and the channel-forming proteins were reconstituted into planar phospholipid membranes as previously described (Rostovtseva & Bezrukov, 2015); briefly, Planar phospholipid bilayers membranes were formed over a hole having a diameter of approximately∼ 100 μm on a 20 μm thick Teflon partition using different lipid mixtures of 1,2-dioleoyl-phosphatidylcholine, di-oleoyl-phosphatidylethanolamine, and dioleoylphosphatidylglycerol. The lipid ratio among DOPC, DOPE, and DOPG was 1:1:2 (w/w). All lipids used in reconstitution were purchased from Avanti Polar Lipids, Inc. (Alabaster, AL). Lipids dissolved in pentane at 5 mg/ml were then carefully added to each chamber. Reconstitution of VDAC proteins was observed after adding ∼5 μL of the porin to the surface of the solution of the cis side of the membrane chamber (the one to which the voltage was applied) concerning the trans-side of the membrane, which was held at the ground. VDAC voltage-gating was obtained on a multichannel lipid bilayer membrane on both sides using a previously described procedure (Colombini, 1989; Queralt-Martín et al., 2019).
Results and Discussion
The voltage dependence has been explained by the movement of charged groups through the entire membrane due to the applied electrical field (Robertson et al., 2021). The net charge on the walls that lines the aqueous pore of VDAC was estimated to be +3.5 for most eukaryotic porins (Zambrowicz & Colombini, 1993). However, for porin from the bovine heart, it was significantly lower, which indicated that this pore-forming protein is less voltage-dependent (Aiche et al., 2022; Fayiah et al., 2022; Pimple, 2022; Vamvuka et al., 2022).
The voltage gating mechanism for VDAC is still not fully understood, though various voltage sensor models exist. Some basic residues of VDAC are present at small enough stoichiometry to permit reliable conclusions to be drawn from specific chemical modifications. The characterization of functionally important amino acid residues by reagents that interact with specific amino acids has provided important clues regarding the mechanism of the VDAC voltage regulation. To further investigate VDAC modulation by covalent modification reagents, the potential involvement and biological relevance of the three histidine residues located at positions 121, 180, and 272 in VDAC activity were proposed. Therefore, histidine residues have been modified to determine if they affect the functional properties of the porin channel (Alshehri et al., 2022; Dirican, 2022).
Treatment of bovine heart mitochondrial porin with DEPC resulted in time-and concentration-dependent modification of histidine and lysine residues. The extent and the time course of carbethoxylation reaction of the porin-histidine residues by DEPC are reflected in the difference absorbance spectra at 242 nm between DEPC-modified and unmodified porin. As illustrated in (Figure 1), indeed, most of the modifying effect of 1.0 mM DEPC on LDAO-porin was reached within the initial 20 minutes of treatment; it reacts primarily with histidine residues but can also react to a lesser extent with lysine residues after 60 minutes of incubation, inactivating mitochondrial porin.
|
|
|
Figure 1. UV differential absorbance spectra of native versus EF-porin at 242 nm. Scans were repeatedly collected over every 10-min period. The reference cuvette contains the LDAO-porin with an equal volume of ethanol used in diethylpyrocarbonate dilutions solution, while the sample cuvette contains the diethylpyrocarbonate-porin solution. |
For the lysine residue to be modified on the epsilon amine group by diethylpyrocarbonate, it must be in its neutral, deprotonated, reactive form. The slow process of the lysine residues of porin with DEPC is probably due to the high pKa values of these lysine residues, making this reaction slow (Pahari et al., 2019). However, the fact that DEPC can modify lysine residues at pH 7.2 indicates that the lysines with the lowest pKa are the kinetically favored residues at slightly basic pH. Incubation of aliquots of carbethoxylated porin with 0.33 M hydroxylamine for 30 min at 4°C that resulted in the absence of any difference in spectra at 242 nm indicates almost complete removal of the ethoxyformyl-group from the histidine residues while leaving the ethoxyformyl group still attached to the ε-amino group of lysine residues as shown in Figure 2.
The reactivation of porin further confirmed the relative reactivity of lysine residues in VDAC after 18 hrs of hydroxylamine treatment. Reversal of DEPC modification by hydroxylamine demonstrates two points. First, at low concentrations of diethylpyrocarbonate, bis-modification of histidine residues with two DEPC molecules apparently could not take place because a bis-carbethoxylation of histidine residues is irreversible. Thus, the inactivation of EF-porin and the parallel decrease in the absorbance at 242 nm indicates possible involvement of the lysine side chain. Second, the reversibility of porin modification and restoration of EF-porin fraction activity can be used as a test to identify the regulatory and functional importance of modified lysine and histidine residues. This would be true, particularly when associated with a hyper-reactive lysine residue.
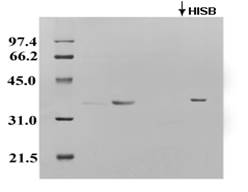
The side chains of histidine and lysine residues in the modified porin are most likely to be exposed on the protein's surface. Therefore, to determine the active regions of porin directly, the LDAO-solubilized porin was incubated with different concentrations of DEPC, and the resulting modified protein was loaded onto a cation exchanger (CMC-52) chromatography column. The column partially retained the protein, and the remaining protein was eluted from the cation resin with a higher ionic strength eluting buffer (Figure 2).
|
|
|
Figure 2. SDS-PAGE of the EF-Porin eluted from the CMC-52 column. First, EF-porin samples were applied to the CMC-52 column and eluted with the equilibration buffer; early eluting fractions were collected, and then elution was achieved by a high ionic strength buffer (HISB). Finally, aliquots of the resulting samples were analyzed by SDS-PAGE. |
These intermediates appear after about a 30-minutes treatment with 20 mM DEPC. Thus, the protein can be modified slowly, partially diminishing its ability to bind to CM-52. In addition, the porin binding capability can be restored after DEPC modification by treating the sample with 0.33 M hydroxylamine at pH 7.5 for 18 hrs.
In the absence of specific covalent reagents, the column retained porin, and elution was most commonly achieved by gradually increasing the ionic strength of the eluting buffer. This approach has been used to identify the gating charges of porin (Aljamal, 2002).
Kinetic analysis of the voltage-dependence gating of the channels before and after the removal of the carbethoxy modifying group by hydroxylamine was performed as described previously (Colombini, 1989; Queralt-Martín et al., 2019). Quantifying the porin gating voltage dependence was determined by applying various potentials to the membrane and measuring the resulting current. Incubation of the retained fraction of EF-porin obtained from experiments performed at 20 mM DEPC concentration with NH2OH and subsequent measurements before and after does not alter the activity of the VDAC. In contrast, the unretained EF-porin fraction significantly increased the voltage dependence by decreasing the optimal probability of finding the open channel. Following the decay in current with an applied potential of 10 mV, the potential was reduced to 5.0 mV and returned to 10 mV, allowing the closed channels to reopen. The closure seen at 10 mV would have required an applied potential of at least 60 mV in the absence of the DEPC. Thus, carbethoxylation of the histidine residues is not responsible for the observed increase in the voltage dependence of channel activation (Alharbi et al., 2022; Blewi et al., 2022).
The ion selectivity and single channel properties were tested to see if ethoxyformylation affected the ions' permeation through the channel. Further information about EF porin channel property and its influence on ion selectivity was obtained from Zero-current potential measurements in the presence of salt gradients across lipid bilayer membranes Variation of VDAC's reversal potentials obtained for the unmodified and EF-porin were, respectively, +7 mV and -6 mV performed in the presence of a 1.0 M vs. 0.1 M KCl solution. The signs of potentials refer to the other side, and the positive sign corresponds to the high salt side (the cis side) negative regarding the low salt side defined as the trans side, i.e., selectivity for K+ over Cl-. After reversing the lysine porin modification, the measured reversal potential (zero-current potential) was +4 mV.
The single channel conductance of the EF-porin obtained from experiments performed at a high DEPC concentration is smaller than the unmodified porin; such a decrease in the single channel conductance was found to be similar to those shown at much higher voltages in the absence of the DEPC (Figure 3).
|
|
|
a) |
|
|
|
b) |
|
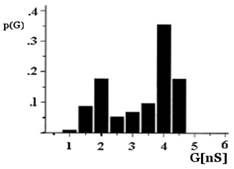
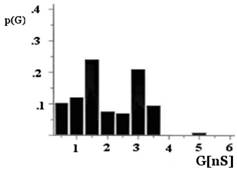
Figure 3. Histogram of conductance fluctuations of porin and EF-porin. The applied potential was +10 mV; T = 20 °C. Representation of the single channel conductance recordings peaks obtained with the EF-porin had two maxima at 1.5 and 3 nS (b) instead of 2 and 4 nS for the porin (a). p(G), is the probability that the single channel conductance (G) is observed. |
The single channel analysis of conductance of the mitochondrial porin from the bovine heart exhibits a broad distribution with a mean of approximately 4 nS (Figure 3a). The EF-porin also shows a broad distribution for the single channel conductance values, but the mean value decreases to 3 nS (Figure 3b). However, no significant difference was detectable at 1.0 mM DEPC concentration, and the size of the conductance fluctuations was equal to those of the control experiments.
Thus, although the relative ion permeability properties of the channel were affected by the chemical modification of the pore-forming protein induced by high concentrations of DEPC, there were no significant changes in either the single channel conductance or the channel anion selectivity at low DEPC concentrations.
Conclusion
Histidine residues in a nucleophilic environment were initially considered candidate residues involved in the voltage-gating mechanisms. However, this possibility is now excluded since there were no functional consequences of modification of histidine residues on the mitochondrial porin activity. It appears reasonable to conclude, therefore, that the observed decrease in the activity of EF-porin at high DEPC concentrations was due to lysine residues modification rather than histidine residues, suggesting that lysine residues are part of the voltage gating mechanism.
Acknowledgments: This research was supported by the deanship of scientific research, Jerash University.
Conflict of interest: None
Financial support: This research was supported by the deanship of scientific research, Jerash University.
Ethics statement: None
Aiche, S., Smail, F., Chikhaoui, M., & Abdelhadi, S. A. (2022). Influence of age, parity, and fetuses’ number on blood parameters in Rembi ewes during late pregnancy in semi-arid zone, Algeria. World Journal of Environmental Biosciences, 11(2), 26-33. doi:10.51847/jD3o3GsDHr
Alharbi, M. G. T., AlHadhari, Y. A. O., AlAnazi, N. F. G., Alanazi, R. N., Alanazi, H. O., AlAnazi, A. S. B., AlJohani, A. M. S., Alanazi, A. S. B., Alruwaili, W. A. O., Alanazi, B. A. Q., et al. (2022). Awareness of obstructive lung diseases and its risk factors, a systematic review. World Journal of Environmental Biosciences, 11(2), 54-60. doi:10.51847/vJAxniXLu3
Aljamal, J. A. (2002). Characterization of different reactive lysines in bovine heart mitochondrial porin. Biological Chemistry, 383(12), 1967-1970. doi:10.1515/BC.2002.222
Alshehri, F. S., Alotaibi, F. F., Alghanim, N. S., Almutairi, F. T., Alsuwailem, H. S., Darwish, E. G., Alharbi, A. A., Alshehri, A. S., Alsubhi, A. S. R., Alfaer, T. A., et al. (2022). Status epilepticus diagnostic and management approach in emergency department. World Journal of Environmental Biosciences, 11(1), 30-33. doi:10.51847/OsSd2wuQQY
Ambasht, P. K. (2020). Use of group-specific reagents in active site functional group elucidation II: Asp, Glu, Arg, Lys, and His residues. In: Singh, D., Tripathi, T. (eds) Frontiers in Protein Structure, Function, and Dynamics. Springer, Singapore. doi:10.1007/978-981-15-5530-5_5
Benz, R. (2021). Historical perspective of pore-forming activity studies of voltage-dependent anion channel (eukaryotic or mitochondrial porin) since its discovery in the 70th of the last century. Frontiers in Physiology, 12, 734226. doi:10.3389/fphys.2021.734226
Biehn, S. E., Limpikirati, P., Vachet, R. W., & Lindert, S. (2021). Utilization of hydrophobic microenvironment sensitivity in diethylpyrocarbonate labeling for protein structure prediction. Analytical Chemistry, 93(23), 8188-8195. doi:10.1021/acs.analchem.1c00395
Blewi, S. M. A., Abuhamad, R. N., Abufara, F. A. I., Bedaiwi, L. A., Alotaibi, J. T. A., Alyami, M. A. M., Alshammari, Y. S., Althobaiti, A. A. H., & Alzahrani, A. M. M. (2022). Epidemiology of epilepsy among pediatric patients in Tabuk City. World Journal of Environmental Biosciences, 11(2), 48-53. doi:10.51847/llGwvXZPYM
Bohm, R., Amodeo, G. F., Murlidaran, S., Chavali, S., Wagner, G., Winterhalter, M., Brannigan, G., & Hiller, S. (2020). The structural basis for low conductance in the membrane protein VDAC upon beta-NADH binding and voltage gating. Structure, 28(2), 206-214. doi:10.1016/j.str.2019.11.015
Colombini, M. (1989). Voltage gating in the mitochondrial channel, VDAC. The Journal of Membrane Biology, 111(2), 103-111. doi:10.1007/BF01871775
Colombini, M. (2016). The VDAC channel: Molecular basis for selectivity. Biochimica et Biophysica Acta. Molecular Cell Research, 1863(10), 2498-2502. doi:10.1016/j.bbamcr.2016.01.019
Diederichs, K. A., Buchanan, S. K., & Botos, I. (2021). Building better barrels – β-barrel biogenesis and insertion in bacteria and mitochondria. Journal of Molecular Biology, 433(16), 166894. doi:10.1016/j.jmb.2021.166894
Dirican, S. (2022). A look at the change in water occupancy rates of gölova dam lake, Turkey. World Journal of Environmental Biosciences, 11(1), 34-36. doi:10.51847/3u8KMQDDzQ.
Ellenrieder, L., Dieterle, M. P., Doan, K. N., Mårtensson, C. U., Floerchinger, A., Campo, M. L., Pfanner, N., & Becker, T. (2019). Dual Role of Mitochondrial Porin in Metabolite Transport across the Outer Membrane and Protein Transfer to the Inner Membrane. Molecular Cell, 73(5), 1056-1065. doi:10.1016/j.molcel.2018.12.014
Endo, T., & Sakaue, H. (2019). Multifaceted roles of porin in mitochondrial protein and lipid transport. Biochemical Society Transactions, 47(5), 1269-1277. doi:10.1042/BST20190153
Fayiah, M., Tulcan, R. X. S., Bockarie, R., Saccoh, S., Singh, S., & Fayiah, M. S. (2022). Sierra Leone rainfall and drought dynamics from 1990-2020: a holistic perspective. World Journal of Environmental Biosciences, 11(2), 34-42. doi:10.51847/8Wz28ID8Mn
Gattin Z, Schneider R, Laukat Y, Giller K, Maier E, Zweckstetter M, Griesinger C, Benz R, Becker S, & Lange A. (2015). Solid-state NMR, electrophysiology, and molecular dynamics characterization of human VDAC2. Journal of Biomolecular NMR, 61(3-4), 311-320. doi:10.1007/s10858-014-9876-5
Höhr, A. I. C., Lindau, C., Wirth, C., Qiu, J., Stroud, D. A., Kutik, S., Guiard, B., Hunte, C., Becker, T., Pfanner, N., et al. (2018). Membrane protein insertion through a mitochondrial beta-barrel gate. Science, 359(6373), 289. doi:10.1126/science.aah6834
Hunsicker-Wang, L. M., & Konkle, M. E. (2018). Translation of chemical biology research into the biochemistry laboratory: Chemical modification of proteins by diethylpyrocarbonate, Best Practices for Supporting and Expanding Undergraduate Research in Chemistry (Vol. 1275, pp. 165-179). Washington, DC: American Chemical Society. doi:10.1021/bk-2018-1275.ch010
Jaremko, M., Jaremko, L., Villinger, S., Schmidt, C. D., Griesinger, C., Becker, S., & Zweckstetter, M. (2016). High-resolution NMR determination of the dynamic structure of membrane proteins. Angewandte Chemie (International ed. in English), 55, 10518-10521. doi:10.1002/anie.201602639
Kayser, H., Kratzin, H. D., Thinnes, F. P., Götz, H., Schmidt, W. E., Eckart, K., & Hilschmann, N. (1989). Zur kenntnis der porine des menschen II. Charakterisierung und primärstruktur eines 31-kDa-porins aus menschlichen B-lymphozyten (porin 31HL). Biological Chemistry Hoppe-Seyler, 370, 1265-1278.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227(5259), 680-685. doi:10.1038/227680a0
Martfeld, A. N., Greathouse, D. V., & Koeppe, R. E. (2016). Ionization properties of histidine residues in the lipid bilayer membrane environment. The Journal of Biological Chemistry, 291(36), 19146-19156. doi:10.1074/jbc.M116.738583
Martynowycz, M. W., Khan, F., Hattne, J., Abramson, J., & Gonen, T. (2020). MicroED structure of lipid-embedded mammalian mitochondrial voltage-dependent anion channel. Proceedings of the National Academy of Sciences of the United States of America, 117(51), 32380-32385. doi:10.1073/pnas.2020010117
Najbauer, E. E., Becker, S., Giller, K., Zweckstetter, M., Lange, A., Steinem, C., de Groot, B. L., Griesinger, C., & Andreas, L. B. (2021). Structure, gating, and interactions of the voltage-dependent anion channel. European Biophysics Journal, 50(2), 159-172. doi:10.1007/s00249-021-01515-7
Najbauer, E. E., Tekwani Movellan, K., Giller, K., Benz, R., Becker, S., Griesinger, C., & Andreas, L. B. (2022). Structure and gating behavior of the human integral membrane protein VDAC1 in a lipid bilayer. Journal of the American Chemical Society, 144, 2953-2967. doi:10.1021/jacs.1c09848
Noskov, S. Y., Rostovtseva, T. K., Chamberlin, A. C., Teijido, O., Jiang, W., & Bezrukov, S. M. (2016). Current state of theoretical and experimental studies of the voltage-dependent anion channel (VDAC). Biochimica et Biophysica Acta, 1858(7 Pt B), 1778-1790. doi:10.1016/j.bbamem.2016.02.026
Pahari, S., Sun, L., & Alexov, E. (2019). PKAD: a database of experimentally measured pKa values of ionizable groups in proteins. Database: The Journal of Biological Databases and Curation, 2019, baz024. doi:10.1093/database/baz024
Pimple, N. S. (2022). Virtual population analysis and recruitment pattern of Osteobrama vigorsii (Sykes, 1839) from Nira River, Bhor Maharashtra. World Journal of Environmental Biosciences, 11(1), 53-59. doi:10.51847/QslcwlezoR
Preto, J., & Krimm, I. (2021). The intrinsically disordered N-terminus of the voltage-dependent anion channel. PLoS Computational Biology, 17(2), e1008750. doi:10.1371/journal.pcbi.1008750
Queralt-Martín, M., Bergdoll, L., Jacobs, D., Bezrukov, S. M., Abramson, J., & Rostovtseva, T. K. (2019). Assessing the role of residue E73 and lipid headgroup charge in VDAC1 voltage gating. Biochimica et Biophysica Acta. Bioenergetics, 1860(1), 22-29. doi:10.1016/j.bbabio.2018.11.001
Rappaport, S. M., Teijido, O., Hoogerheide, D. P., Rostovtseva, T. K., Berezhkovskii, A. M., & Bezrukov, S. M. (2015). Conductance hysteresis in the voltage-dependent anion channel. European Biophysics Journal, 44(6), 465-472. doi:10.1007/s00249-015-1049-2
Reif, M. M., Fischer, M., Fredriksson, K., Hagn, F., & Zacharias, M. (2019). The N-Terminal Segment of the voltage-dependent anion channel: A possible membrane-bound intermediate in pore unbinding. Journal of Molecular Biology, 431(2), 223-243. doi:10.1016/j.jmb.2018.09.015
Robertson, J., Ghimire, M. L., & Reiner, J. E. (2021). Nanopore sensing: A physical-chemical approach. Biochimica et Biophysica Acta Biomembranes, 1863(9), 183644. doi:10.1016/j.bbamem.2021.183644
Rostovtseva, T. K., & Bezrukov, S. M. (2015). Function and regulation of mitochondrial voltage-dependent anion channel. In: Delcour, A.H. (eds) Electrophysiology of Unconventional Channels and Pores. Springer Series in Biophysics, vol 18. pp. 3-31, Springer, Cham. doi:10.1007/978-3-319-20149-8_1
Thomas, L., Blachly-Dyson, E., Colombini, M., & Forte, M. (1993). Mapping of residues forming the voltage sensor of the voltage-dependent anion-selective channel. Proceedings of the National Academy of Sciences of the USA, 90(12), 5446-5449. doi:10.1073/pnas.90.12.5446
Vamvuka, D., Teftiki, A., & Sfakiotakis, S. (2022). Investigating the valorisation of refused derived fuel for energetic uses through its co-gasification with woody wastes. World Journal of Environmental Biosciences, 11(1), 37-44. doi:10.51847/2fiOEjSU7L
Zachariae, U., Schneider, R., Briones, R., Gattin, Z., Demers, J. P., Giller, K, Maier, E., Zweckstetter, M., Griesinger, C., Becker, S., et al. (2012). Beta-Barrel mobility underlies closure of the voltage-dependent anion channel. Structure, 20(9), 1540-1549. doi:10.1016/j.str.2012.06.015
Zambrowicz, E. B., & Colombini, M. (1993). Zero-current potentials in a large membrane channel: a simple theory accounts for complex behavior. Biophysical Journal, 65(3), 1093-1100. doi:10.1016/S0006-3495(93)81148-2
Zeth, K., & Zachariae, U. (2018). Ten years of high-resolution structural research on the voltage-dependent Anion Channel (VDAC)-recent developments and future directions. Frontiers in Physiology, 9, 108. doi:10.3389/fphys.2018.00108
Zhou, Y., & Vachet, R. W. (2012). Diethylpyrocarbonate labeling for the structural analysis of proteins: label scrambling in solution and how to avoid it. Journal of the American Society for Mass Spectrometry, 23(5), 899-907. doi:10.1007/s13361-012-0349-3