Diversity and Density of Fungi Isolated from Dried Fruits
|
Rana G. Alghamdi, Nidal M. Zabermawi, Fardos M. Bokhari, Rania M. Makki, Shahira A. Hassoubah, Zeina W. Sharawi, Azhar A. Najjar* Department of Biological Sciences, Faculty of Sciences, King Abdulaziz University, Jeddah 21473, Saudi Arabia.
Faizah A. Altihani Department of Biology, Faculty of Sciences, King Khalid University, Abha 61413, Saudi Arabia.
|
*E-mail: [email protected]
Abstract
Fruits are a vital source of nutrients for human health, containing carbohydrates, minerals, vitamins, and amino acids. However, their nutritional value also makes them favorable for the growth of microorganisms. Dried fruits, can be susceptible to fungal contamination, which can pose significant health risks. In a study conducted in Jeddah, Saudi Arabia, a total of 180 imported and local dried fruit samples, including raisins, dates, and apricot, were purchased from local markets. The study involved fungal isolation, where each fruit was cut into small segments (5x5 mm) and cultured on potato dextrose agar. The samples were then incubated at 25º C for 7-14 days to identify phenotypic fungal characteristics. Also, molecular analysis of the fungal isolates was performed by sequencing the internal transcribed spacer (ITS) region of the rRNA gene. The morphological and molecular combination results of the study revealed the presence of 17 different fungal species belonging to 13 genera, including Alternaria, Aspergillus, Cladosporium, Curvularia, Eurotium, Lecythophora, Mucor, Paecilomyces, Penicillium, Rhizopus, Talaromyces, Ulocladium, and Zygosaccharomyces. Among the dried fruits, the most dominant genera isolated from raisins and dates were Aspergillus sp. (367 CFU), Penicillium sp. (74 CFU), and Rhizopus sp. (27 CFU). In dried apricots, Zygosaccharomyces sp. (2932 CFU) was the only detected isolated fungus. Additionally, a very toxic effect of the selected fungal extracts on brine shrimp larvae was noted. In conclusion, it is crucial to implement good manufacturing practices (GMPs) to ensure the production of healthy dried fruits and minimize contamination risks during and after harvest.
Keywords: Dried fruits, Fungi, Aspergillus, ITS rRNA, Toxicity
Introduction
Food is an essential source of nutrition for humans, it also acts as a conducive environment for the growth of microbes, some of which may be harmful (Siserman et al., 2019; Galea-Holhoș et al., 2023; Govindaraj et al., 2023). Food safety is a major global issue concerning the production and processing of food (Gherman et al., 2019; Nicoară et al., 2023). A crucial international concern related to food production and processing is food safety (Delcea & Siserman, 2020; Rus et al., 2020; Alhussain et al., 2022). Foodborne illnesses are a serious global public health concern in developing nations with insufficient food, safety policies, and regulations of food (Sousa, 2008). Due to their high nutritional value, which helps prevent illnesses like cardiovascular disease, cancer, and eye disease, fruits, and vegetables are essential for maintaining a healthy body (Pavithra et al., 2023; Radu et al., 2023; Voiţă-Mekereş et al., 2023). Thus, it becomes essential to ensure the safety of fruits and vegetables and implement appropriate measures to reduce the risk of microbial contamination (Redzuan et al., 2023). Fruits and vegetables are a vital component of a diet that is both balanced and healthful in all communities on the planet. Moreover, infections from human, animal, or environmental sources can contaminate fruits and vegetables during growing, harvesting, transportation, processing, and handling—even though they are known to naturally carry nonpathogenic epiphytic microbiota. Numerous outbreaks of foodborne illness in the US, Europe, Canada, and Japan have been linked to fresh fruit (Mostafidi et al., 2020). Food commodities are spoiled by both biotic and abiotic factors. Biotic agents especially microorganisms are vital to the deterioration of the nutritive value of stored food commodities amongst whom fungi play an important role during storage. Contamination of dried fruits may indicate inadequate storage and drying procedures. Sufficiently dried fruits are extremely low in moisture and include microbes such as fungi and bacteria (Dsouza, 2021). As previously noted in organic raisin samples, several dried fruits may contain bacterial isolates, including coliform bacteria like E. coli. Mango and raisin samples included fungi, including the mold Aspergillus (Torres, 2007; Alp & Bulantekin, 2021).
High levels of sugars, optimal water activity, harvesting techniques, and drying conditions are the main triggers of fungal contamination in dried fruits, which might give rise to mycotoxin development and fungus infection (Ali et al., 2020). Fungi-induced fruit contamination might point to inadequate drying, processing, and storage. Due to their high ability to produce aflatoxins, A. flavus and A. parasiticus are the most common fungal species contaminating food including dried fruits. These two aflatoxigenic species are cosmopolitan and may grow and form aflatoxins under many conditions in a wide range of environments (Hocking et al., 1997). Microorganisms’ numbers on most dried fruits ranged from a few hundred per gram of fruits to thousands majority of them being on the outside surfaces (Kallagova et al., 2023). In cases when the drying trays are dirty and improperly packed, resulting in spoiling, there is a discernible rise in the number of bacteria and fungus during the drying process. The spoilage occurs in most dry fruits during handling and storage. A significant quantity of mold spores may exist when part of the fruit has supported the growth and sporulation of the mold before or after drying (Zhao et al., 2022). These microbes can cause diseases based on their low infectious dose or favorable temperature that allows the organism to grow (Lauer et al., 2021).
Numerous fungi can produce mycotoxins, including ochratoxin A and aflatoxin B1. The eating of dried fruits or juices and jams manufactured with the dried fruits exposes humans to mycotoxins (Voiţă-Mekereş et al., 2023). To prevent mycotoxins and microflora, which are harmful to human health, from contaminating dried fruits, stringent microbiological hygiene procedures must be followed at all times, including during harvest, handling, transportation, storage, and drying. Foodborne disease occurs when dried fruits come into interaction with vegetative pathogens or their toxins, mostly from animal products. Consumption of contaminated food is the major pathogen impacting food safety and causing human diseases globally in people all over the world (Alghalibi & Shater, 2004; Özer et al., 2022).
In this study, fungi were isolated from a pool of 180 dried fruits, comprising both imported varieties and locally sourced ones. These fruits were collected from diverse markets in Jeddah. The identification of fungal isolates involved morphological and molecular methods. Additionally, a screening for toxicity activity was conducted on selected fungal isolates to assess their capability to produce toxic metabolites.
Collection of Dried Fruits Samples
A total of 180 dried fruit samples including Raisins, Dates, and apricots, encompassing both imported and local varieties, were collected under normal purchase conditions in January 2020 from different markets in Jeddah Governorate, Saudi Arabia. These samples were carefully placed in sterilized polyethylene bags and transported to the laboratory for the subsequent isolation of fungi.
Isolation of Fungi from Dried Fruits
The direct plating method was used for the fungal isolation process from dried fruit samples. Each of the 60 dried fruit samples (comprising 30 imported and 30 local dried fruits) underwent segmentation into small pieces measuring 5x5 mm² at a rate of four particles per plate, with the process carried out in triplicate. These plates were then kept in incubation at 25oC for 7 days. Subsequently, a single plug (5-mm diameter disk) from each fungal isolate was transferred to the PDA medium, serving as the subculture medium, and incubated for 7–14 days at 25 °C to observe their growth and sporulation. Macro-morphological characteristics including colony color, radial growth (mm), and pigment production were documented.
Diversity and Density Analysis
Pure cultures grown on PDA media were analyzed for the phenotypic identification of all fungal isolates. Important diagnostic characteristics for identification as colony color and texture, growth rate, and reverse pigmentation were carried out. The microscopic features of the growing fungi hyphae and spores were carefully observed in wet mounts made on microscopic slides and stained with lactophenol cotton blue (LPCB). To determine the number of fungal isolates, colony forming units (CFU) of each fungal species were calculated in density studies using the method described before by Costa et al. (2012).
Molecular Identification of Fungal Isolation
The morphological identification of fungal isolates was done based on their macro and micro characteristics. Then the fungal isolates were subsequently identified using sequences of the ITS region of the ribosomal RNA (ITS rRNA) (Al-Jaradi et al., 2018). DNA extraction followed the method outlined by Al‐Samarrai and Schmid (2000) with slight adjustments. Fungal isolates were inoculated in 100 ml of Erlenmeyer flasks containing 20 ml of potato dextrose broth (PDB) and incubated at 28°C for 5 days. Subsequently, mycelia were filtered using sterilized filters. The fungal mycelium was finely powdered using a liquid nitrogen-sanitized. The resulting fine powders were transferred to a 1.5 ml Eppendorf tube and stored at -20°C. For DNA extraction, 30 mg of frozen ground mycelium were resuspended and lysed in 500 μl of lysis buffer (pH 7.8) and incubated in a water bath at 37°C for 60 mins. The lysis buffer comprised 1% SDS, 20 mmol/1 sodium acetate, 40 mmol Tris-acetate, and 1 mmol EDTA. The DNA was extracted by using a Gene JET Genomic DNA extraction kit (Thermo Scientific, USA). The internal transcribed spacer (ITS) regions of fungal ribosomal DNA were amplified using two primers; ITS1 (5′CTTGGTCATTTAGGGAAGTAA3′) and ITS4 (5′TCCTCCGCTTATTGATATGC3′) in a thermal cycler (Esco health care, Swift max pro, Malaysia) programmed as follows: 94°C for 4 min, 32 cycle loop start, 94°C for 1 min, 51°C for 1 min, 72°C for 1 min, cycles end, finally samples were incubated as final extension at 72°C for 10 min. The reaction mixture (50 μl) included 3 μl of templet DNA, 5 μl each of the primers, 25 μl of green PCR mix (Promega, Go Taq® Green Master Mix, USA), and 50 μl of nuclease-free water (Najjar et al., 2020). The PCR products were loaded onto 1.5% agarose gel and electrophoresed for 45 minutes at 130 volts using Horizontal gel electrophoresis equipment (Cleaver Scientific, UK). The gel was stained using ethidium bromide and visualized under UV light using a gel documentation imager (Biorad. USA). To quantify and identify PCR products, a 100 bp DNA Ladder (Invitrogen, USA) was used as a DNA marker. Then the samples underwent purification and sequencing at Macrogen company in South Korea. Manual trimming of low-quality bases was performed, retaining only the highest-quality bases for analysis. The DNA sequences of the fungal isolates were subjected to BLAST analysis (blast.ncbi.nlm.nih.gov) using default parameters with sequencing data compared against the nucleotide collection database.
Toxicity Testing of Fungal Extracts
To conduct toxicity testing, brine shrimp larvae Artemia salina (5g) were hatched in a 300 ml Erlenmeyer flask containing 100 ml of natural seawater and incubated at 28°C on a rotary shaker (operating at 140 revs min-1) for 24-30h, under light aeration. After hatching, larvae were collected into fresh seawater at room temperature. Various dilutions of fungal extracts were prepared in DMSO at concentrations of 20, 40, 60, and 80 µg/ml of fungal extracts in small dishes, with each dilution tested in triplicate. Ten A. salina larvae were added to each dish, and the final solution volume in each dish was adjusted to 7 ml with seawater immediately after adding the shrimps. The dishes were then incubated at room temperature. After 6 hours, the average number of larvae that survived in each dish was determined (Adoum, 2009, Momoh et al., 2021). The count of survivors was recorded and the percentage of deaths was calculated. Larvae were considered dead if they showed no internal or external movement during several seconds of observation (Idris et al., 2019). The toxicity of extracts was rated using the method of Hartwig as follows: 0-9% mortality- non-toxic (NT), 10-49% mortality- slightly toxic (ST), 50-89% mortality- toxic (T), and 90-100% mortality- very toxic (VT).
Results and Discussion
Morphological Analysis of Fungal Isolates
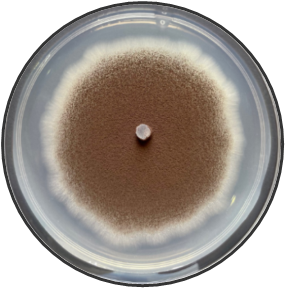
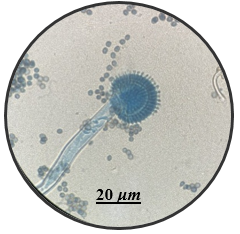
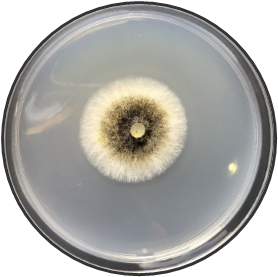
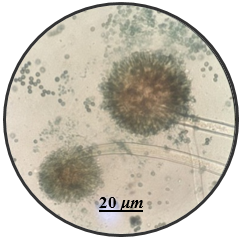
In this study, the aim was to investigate the diversity and density of fungi in 180 dried fruit samples, including imported and local comprising raisins, dates, and apricots, Saudi Arabia. A total of 17 fungal species belonging to 13 genera were isolated and identified. These fungi belonged to the genera Alternaria, Aspergillus, Cladosporium, Curvularia, Eurotium, Lecythophora, Mucor, Paecilomyces, Penicillium, Rhizopus, Talaromyces, Ulocladium, Zygosaccharomyces as shown in Table 1. Morphological reference keys, as described by Tsuneo (2010), were used to record morphological features of fungi at the genus level. Characteristics such as colony color and surface properties were documented by observing the pure cultures on PDA medium, which were incubated at 25°C for 7-14 days to detect the distinct conidial features. Detailed descriptions along with macro and microscopic images of the most prevalent fungal isolates including Aspergillus spp., Penicillium spp., and Zygosaccharomyces sp. were included in Figure 1. The total count of isolated fungi amounted to 3417 isolates from various imported and local dried fruit samples specifically raisins, dates, and apricot samples. The most prevalent genera identified in two types of dried fruits namely raisins and dates were Aspergillus sp. (367 CFU), Penicillium sp. (74 CFU), Rhizopus sp. (27 CFU), Lecythophora sp. (7 CFU), Alternaria sp. (2 CFU), Curvularia sp. (2 CFU), Cladosporium sp. (1 CFU), Eurotium sp. (1 CFU), Mucor sp. (1 CFU), Paecilomyces sp. (1 CFU), Talaromyces sp. (1 CFU), Ulocladium sp. (1 CFU). In dried apricot Zygosaccharomyces sp. (2932 CFU) was the only detected isolated fungi as shown in Table 1. Notably, the total isolated fungi of imported dried fruit samples were more than the total isolated fungi of local dried fruits. Previous studies in Jeddah reported similar fungal genera such as Alternaria, Aspergillus, Penicillium, Rhizopus, Mucor, and Paecilomyces in dried fruits (Al Ghamdi et al., 2019). Some of these species are also common in commercially imported dried fruits from Iraq and South Africa (Saadullah & Abdullah, 2014; Ramadan et al., 2022), where the presence of pathogenic fungi and toxins has been documented by Boyacioglu and Gönül (1990). Analysis of raisins identified A. niger as the most common contaminant, with A. flavus being less frequently isolated in this study. Similar findings were reported in the Yemen Republic, where A. niger prevalence on dry raisins was noted, and A. flavus was less common (Alghalibi & Shater, 2004; Özer et al., 2022). Gherbawy et al. (2012) conducted a study on the mycological characteristics of commercially available date fruits in various markets in Taif, Saudi Arabia. From 50 dates samples, they isolated 22 fungi belonging to 12 genera. The most prevalent fungi included A. flavus, A. niger, P. chrysogenum, and Rhizopus stolonifera. Moreover, Toma and Rajab (2014) analyzed fungi on dried fruits in Iraq, revealing the presence of 15 genera and 16 species of various fungi. The most dominant species were 16 species of Aspergillus, 5 species of Penicillium, 2 species of Alternaria, 2 species of Gliocladium, 4 species of Mucor, and 5 species of Rhizopus. Additional isolated fungi comprised Emericella nidulans, Geotrichum sp., Helminthosporium australians, Mortierella sp., Neurospora sp., Paecilomyces victoria, Rhizoctonia sp., and Scopularisopsis sp. were isolated in lower incidence rate. According to Victor et al. (2017), dried fruits are susceptible to various fungi, known to pose significant threats to fruit production and the quality of dried fruits. Spoilage defined as any alteration in food quality leading to unpalatability or even toxicity, is a prevalent concern. In addition, a report from Nigeria by Colman et al. (2012) suggested that date palm fruits are more prone to contamination with Aspergillus spp. than other fungal species. In their study, among the six fungal species isolated, A. niger had the highest occurrence rate (39.17%), followed by A. flavus (17.60%), Mucor sp. (16.67%), A. fumigatus (12.50%), and Trichophyton rubrum (4.16%), with Candida albicans having the lowest percentage of occurrences (0.83%). Hakobyan et al. (2010), stated that the mycological examination of 41 Armenian raisins samples and 11 foreign raisins samples gathered from various Yerevan marketplaces revealed a high prevalence of Aspergillus species, constituting 65.2% of the total frequency. Thirty-two filamentous fungus species belonging to Aspergillus, Penicillium, Alternaria, Trichoderma, and Syncephalastrum genera were isolated and identified.
Drying, among the earliest preservation methods, can be applied to fruits in various forms, sliced, or pureed. Some fruits such as grapes are typically dried whole; bananas, mango, papaya, and kiwi are sliced; while mango and apricot are dried in puree form. The reduction of water content during drying serves to prevent microbial growth and enzymatic reactions, making the fruits resistant to spoilage. Low moisture, high acidity, sugar, and the drying process collectively contribute to this resistance. Microbial populations on dried fruits vary from a few hundred to thousands per gram, predominantly on the outer surfaces. Mold spores become significant when a part of the fruit supports mold growth and sporulation before or after drying. Improper loading and cleaning of drying trays can lead to increased fungi quantities during the drying process. Handling, storage, and transportation often contribute to the spoilage of most dried fruits. The presence of mycotoxins, known to compromise food quality and pose health risks, is a significant concern for the global public health, agriculture, and economy (Yuvaraj et al., 2023). Exporting countries, in particular, face health hazards and financial losses due to mycotoxin contamination in dried fruits. The low water activity and high sugar concentration found in dried fruits create favorable conditions for the development of Aspergillus species, particularly A. niger, and A. ochraceus, as they are xerophilic. Among these, A. niger was identified as the most prevalent fungus in the majority of dried fruit samples. It is emphasized fungi as the primary culprits behind the spoilage of high-moisture dried fruits (Toma & Rajab, 2014). pointed out that fungi exhibit greater tolerance to reduced water activity compared to bacteria. Unhygienic conditions and unsanitary environments during the preparation of home-dried fruits and vegetables contribute to the presence of pathogenic microorganisms (Kumar & Singh, 2022). For instance, finely slicing fruits and vegetables with bare hands in open settings before drying exposes the products to potential microbial contamination. Additionally, the drying process, conducted openly in the sun for several days, further exposes the items to the risk of contamination (Victor et al., 2017). Fungi necessitate specific environmental conditions for growth and mycotoxin production. Ensuring the safety of fruits presents a significant challenge due to the perceived inevitability and unpredictability of mycotoxin contamination, even with the implementation of ideal agricultural, storage, and processing practices. Consequently, the adoption of appropriate food safety training, such as Hazard Analysis and Critical Control Points (HACCP), Good Manufacturing Practices (GMP), and Good Hygienic Practices (GHP), becomes crucial, as these measures can effectively mitigate pathogenic hazards in food (Ijabadeniyi & Buys, 2012).
Table 1. Diversity and density of fungi obtained from three distinct types of imported and locally sourced dried fruits (raisins, dates, apricots) by direct plating method on PDA medium for 7-14 days at 25°C.
|
Isolate No. |
Dried fruits
Fungal species |
Raisins |
Dates |
Apricot |
Total Count (CFU) |
||||
|
Imported Raisins (Yemeni) |
Local Raisins (Saudi) |
Imported Dates (Egyptian) |
Local Dates (Saudi) |
Imported Apricot (Turkish) |
Local Apricot (Saudi) |
||||
|
Alternaria |
2 |
- |
- |
2 |
|||||
|
YR1 |
Alternaria tenuissima |
2 |
- |
- |
- |
- |
- |
2 |
|
|
Aspergillus |
143 |
224 |
- |
367 |
|||||
|
ED2 |
A. flavus |
- |
- |
11 |
1 |
- |
- |
12 |
|
|
SD3 |
A. japonicas |
- |
- |
- |
1 |
- |
- |
1 |
|
|
YR4 |
A. niger |
16 |
123 |
89 |
122 |
- |
- |
350 |
|
|
YR5 |
A. ustus |
4 |
- |
- |
- |
- |
- |
4 |
|
|
Cladosporium |
1 |
- |
- |
1 |
|||||
|
YR6 |
Cladosporium cladosporioides |
1 |
- |
- |
- |
- |
- |
1 |
|
|
Curvularia |
2 |
- |
- |
2 |
|||||
|
YR7 |
Curvularia spicifera |
2 |
- |
- |
- |
- |
- |
2 |
|
|
1 |
- |
- |
1 |
||||||
|
YR8 |
Eurotium sp. |
1 |
- |
- |
- |
- |
- |
1 |
|
|
Lecythophora |
4 |
3 |
- |
7 |
|||||
|
YR9 |
Lecythophora canina |
4 |
- |
- |
3 |
- |
- |
7 |
|
|
Mucor |
1 |
- |
- |
1 |
|||||
|
SR10 |
M. racemosus |
- |
1 |
- |
- |
- |
- |
1 |
|
|
Paecilomyces |
1 |
- |
- |
1 |
|||||
|
YR11 |
Paecilomyces variotii |
1 |
|
- |
- |
- |
- |
1 |
|
|
Penicillium |
74 |
- |
- |
74 |
|||||
|
YR12 |
P. chrysogenum |
60 |
|
- |
- |
- |
- |
60 |
|
|
YR13 |
P. polonicum |
14 |
|
- |
- |
- |
- |
14 |
|
|
Rhizopus |
24 |
3 |
- |
27 |
|||||
|
YR14 |
Rhizopus sp. |
10 |
14 |
- |
3 |
- |
- |
27 |
|
|
Talaromyces |
- |
1 |
- |
1 |
|||||
|
ED15 |
T. wortmannii |
- |
- |
1 |
- |
- |
- |
1 |
|
|
Ulocladium |
1 |
- |
- |
1 |
|||||
|
YR16 |
Ulocladium sp. |
1 |
|
- |
- |
- |
- |
1 |
|
|
Zygosaccharomyces |
- |
- |
- |
2932 |
|||||
|
TA17 |
Z. rouxii |
- |
- |
- |
- |
- |
- |
2932 |
|
|
Total count in all samples |
|
3417 |
|||||||
(-) = Not detected
The study revealed that the predominant fungal species identified were mainly from the Ascomycota division, comprising 15 out of 17 species. In contrast, only two species, namely Mucor racemosus and Rhizopus sp., were associated with the Zygomycota division. The classification of these fungi into various families, orders, and classes is detailed in Table 2. Typically, the identification of fungi involves a combination of genotypic and phenotypic characterization to establish a comprehensive overview of closely related species. This approach is adopted due to the challenges in microscopically distinguishing certain fungal features.
Table 2. classification of fungal strains isolated from three different types of dried fruits.
|
Genera and species |
Family |
Order |
Class |
Division |
|
Alternaria tenuissima |
Pleosporaceae |
Pleosporales |
Dothideomycetes |
Ascomycota |
|
Aspergillus flavus |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Aspergillus japonicas |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Aspergillus niger |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Aspergillus ustus |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Cladosporium cladosporioides |
Cladosporiaceae |
Capnodiales |
Dothideomycetes |
Ascomycota |
|
Curvularia spicifera |
Pleosporaceae |
Pleosporales |
Dothideomycetes |
Ascomycota |
|
Eurotium sp. |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Lecythophora sp. |
Coniochaetaceae |
Coniochaetales |
Sordariomycetes |
Ascomycota |
|
Mucor racemosus |
Mucoraceae |
Mucorales |
Zygomycetes |
Zygomycota |
|
Paecilomyces variotii |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Penicillium chrysogenum |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Penicillium polonicum |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Rhizopus sp. |
Mucoraceae |
Mucorales |
Zygomycetes |
Zygomycota |
|
Talaromyces wortmannii |
Trichocomaceae |
Eurotiales |
Eurotiomycetes |
Ascomycota |
|
Ulocladium sp. |
Pleosporaceae |
Pleosporales |
Dothideomycetes |
Ascomycota |
|
Zygosaccharomyces rouxii |
Saccharomycetaceae |
Saccharomycetales |
Saccharomycetes |
Ascomycota |
|
|
|
|
a) Aspergillus flavus |
|
|
|
|
|
b) Aspergillus japonicas |
|
|
|
|
|
c) Aspergillus niger |
|
|
|
|
|
d) Penicillium chrysogenum |
|
|
|
|
|
e) Penicillium polonicum |
|
|
|
|
|
f) Zygosaccharomyces rouxii |
|
|
Figure 1. Fungi isolated from imported and local dried fruits including raisins, dates, and apricots on PDA medium for 7 days at 25°C. Detailed descriptions along with macro and microscopic images of the most prevalent fungal isolates including Aspergillus spp. (a-c), Penicillium spp. (d-e) and Zygosaccharomyces sp. (f) using light microscopy at 100X magnification. |
|
Molecular Identification of Isolated Fungi
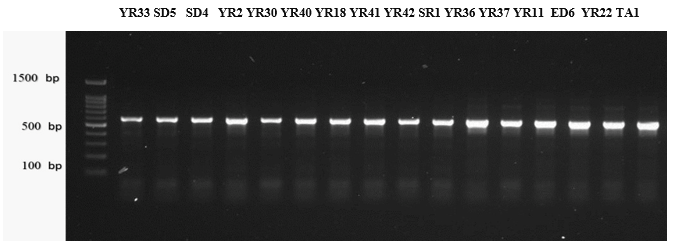
The PCR amplification of DNA extracted from 16 fungal Isolates was utilizing ITS as a universal fungal primer pair. Agarose gel electrophoresis revealed intense bands representing PCR products with expected sizes of around 600 bp for all 16 isolates. The PCR profiles, specifically of the ITS1/4 regions, are visually represented in Figure 2. Sequencing data obtained from these fungal strains were aligned with sequences from closely related strains available in GenBank, and the results are presented in Table 3. Molecular identification of the 16 Strains to the species level was established based on a similarity range 86-100% with sequences of other known species already published in the NCBI database. A dendrogram illustrating the phylogenetic analysis of the sequenced fungal strains is presented in Figure 3. To accurately distinguish and confirm the identities of the isolates at the species level, an advanced molecular technique was employed. This involved amplifying the ITS1 segment for fungal isolates through PCR products and utilizing a sequencing analysis method. Previous studies have attested to the effectiveness of this molecular approach in confirming species identification (Khazamova et al., 2023). The molecular analysis of the microbial strains validated the accuracy of the identification based on morphological features and biochemical tests, ensuring correct identification for all tested isolates (Najjar et al., 2020).
Table 3. Molecular identification of fungal strains, with corresponding GenBank accession numbers and the percentage identity compared to closely related strains.
|
Fungal strains isolated in the current study |
Closely related strains accessed from GenBank |
||||
|
Fungal species |
Strain No. |
Fungal species |
Strain No. |
GenBank No. |
Identity (%) |
|
Alternaria tenuissima |
YR1 |
Alternaria tenuissima |
64APRIL |
97.28% |
|
|
Aspergillus flavus |
ED2 |
Aspergillus flavus |
EF_241 |
99.82% |
|
|
Aspergillus japonicas |
SD3 |
Aspergillus japonicas |
286N |
98.48% |
|
|
Aspergillus niger |
YR4 |
Aspergillus niger |
A7 |
100% |
|
|
Aspergillus ustus |
YR5 |
Aspergillus ustus |
Rn14-81A |
99.61% |
|
|
Cladosporium cladosporioides |
YR6 |
Cladosporium cladosporioides |
LA8 |
100% |
|
|
Curvularia spicifera |
YR7 |
Curvularia spicifera |
Cc-22018 |
86.37% |
|
|
Eurotium sp. |
YR8 |
Eurotium sp. |
72DEC |
97.03% |
|
|
Lecythophora canina |
YR9 |
Lecythophora sp. |
OK-C-AD-4 |
98.31% |
|
|
Mucor racemosus |
SR10 |
Mucor racemosus |
SF_994 |
98.42% |
|
|
Paecilomyces variotii |
YR11 |
Paecilomyces variotii |
RS3-S2-27 |
MN547409.1 |
99.45% |
|
Penicillium chrysogenum |
YR12 |
Penicillium chrysogenum |
CAL1 |
99.80% |
|
|
Penicillium polonicum |
YR13 |
Penicillium polonicum |
sC3 |
98.55% |
|
|
Talaromyces wortmannii |
ED15 |
Talaromyces wortmannii |
UASWS1158 |
96.80% |
|
|
Ulocladium sp. |
YR16 |
Ulocladium sp. |
SR28 |
99.61% |
|
|
Zygosaccharomyces rouxii |
TA17 |
Zygosaccharomyces rouxii |
DMic 124141 |
94.21% |
|
|
|
|
Figure 2. The PCR products from 16 fungal strains were subjected to gel electrophoresis. The strains included : (YR1) Alternaria tenuissima, (ED2) A. Flavus, (SD3) A. japonicas, (YR4) A. Niger, (YR5) A. ustus, (YR6) Cladosporium cladosporioides, (YR7) Curvularia spicifera, (YR8) Eurotium sp., (YR9) Lecythophora canina, (SR10) Mucor racemosus, (YR11) Paecilomyces variotii, (YR12) Penicillium chrysogenum, (YR13) Penicillium polonicum, (ED15) Talaromyces wortmannin, (YR16) Ulocladium sp., and (TA17) Zygosaccharomyces rouxii. The amplification was carried out using the ITS1/4 primer, and the DNA molecular size ranged around 600 bp, with M (100 bp marker) serving as a reference on the left side of the image. |
|
|
|
Figure 3. Dendrogram illustrating polygenetic analysis utilizing the ITS region and referring the NCBI GenBank database for a set of 15 fungal species. |
Toxicity Bioassay
Following a 6-hour observation of the brine shrimp larvae's sensitivity to extracts from selected fungal isolates, the research revealed that all shrimp in the control group survived. A notable increase in mortality rates was observed at the highest concentration range of 60-80 μg/ml, with the lowest mortality occurring at the lowest concentration of 20 μg/ml. Conversely, extracts from A. tenuissima, A. japonicas, T. wortmannii, and Z. rouxii exhibited high lethality across all concentration levels, as indicated in Table 4. It's worth noting that the brine shrimp larvae test, while useful for screening toxic substances, cannot identify the specific toxic metabolites of toxigenic strains. Such bioassay systems serve the purpose of detecting toxic substances, and other compounds like fungicide or pesticide residue, rather than specifically pinpointing mycotoxins. Reiss (1972) utilized A. salina larvae as a model organism to compare the toxicity of 42 fungal secondary metabolites, with 32 of them being mycotoxins. This test is widely used to detect fungal toxins, plant extract toxicity, and other substances. Considered a valuable technique for preliminary toxicity assessment (Solis et al., 1993; Avhad et al., 2023), the A. salina eggs can be easily obtained from pet retailers, developing within 24 to 30 hours, making them a reliable screening system for evaluating the toxicity of fungal extracts. Fungi, along with their toxic byproducts known as mycotoxins, are nearly ubiquitous in food, whether it is fresh or dried fruit, especially in developing regions (Păduraru et al., 2022). Mycotoxins, hazardous secondary metabolites produced by fungi, have the potential to contaminate agricultural goods both before and after harvest. These mycotoxins can pose immediate or delayed harm to both humans and animals, with consequences ranging from osteogenic, mutagenic, carcinogenic, teratogenic to atherogenic effects (Domiaty, 2022). For instance, Alghalibi and Shater (2004) conducted investigations on fungal toxins isolated from various dried fruits in Yemen. They identified 23 species from dried fruits such as raisins, dates, and figs. The most prevalent fungal species, including A. niger, A. flavus, A. fumigatus, A. ochraceus, P. chrysogenum, and R. stolonifer. The study also examined dried fruit samples for various mycotoxins. The brine shrimp bioassay results indicated that over 25% of all samples were toxic to A. salina larvae. Similar findings were reported by Abdel-Sater and Saber (1999) in Egypt, where 30% of dried figs, dates, and raisins samples were toxic to the larvae.
Table 4. Percentage of mortality observed on brine shrimp larvae after treating with the selected fungal extracts.
|
Fungal species |
Mortality after 6 h |
|||
|
μg/mL |
||||
|
20 |
40 |
60 |
80 |
|
|
Alternaria tenuissima |
100% |
100% |
100% |
100% |
|
Aspergillus flavus |
80% |
90% |
100% |
100% |
|
Aspergillus japonicas |
90% |
100% |
100% |
100% |
|
Aspergillus niger |
20% |
40% |
90% |
100% |
|
Aspergillus ustus |
20% |
30% |
100% |
100% |
|
Cladosporium cladosporioides |
30% |
90% |
100% |
100% |
|
Curvularia spicifera |
70% |
100% |
100% |
100% |
|
Eurotium sp. |
20% |
50% |
100% |
100% |
|
Lecythophora canina |
30% |
60% |
90% |
100% |
|
Mucor racemosus |
30% |
100% |
100% |
100% |
|
Paecilomyces variotii |
10% |
20% |
40% |
90% |
|
Penicillium chrysogenum |
10% |
40% |
70% |
100% |
|
Penicillium polonicum |
50% |
100% |
100% |
100% |
|
Talaromyces wortmannii |
100% |
100% |
100% |
100% |
|
Zygosaccharomyces rouxii |
100% |
100% |
100% |
100% |
0-9% mortality - non-toxic (NT)
10-49% mortality- slightly toxic (ST)
50-89% mortality - toxic (T)
90-100% mortality - very toxic (VI)
Conclusion
This investigation assessed the fungal quality of both imported and local dried fruits, in Saudi Arabia, revealing the presence of potentially harmful fungi like Aspergillus, Eurotium, Penicillium and Alternaria and others. The existence of potential patho-toxigenic species on dried fruits poses a considerable public health risk for consumers. There's a risk that plant pathogenic organisms from the field might survive the drying process and end up in the fruits after harvest. It was evident that fruits sold in markets lacked proper covering from airborne contaminants and dust, potentially exposing customers to mycotic illnesses upon consumption. To maintain the quality of these products, it is essential to store them under conditions that sustain appropriate moisture levels throughout storage and marketing.
Acknowledgments: This project was supported by the Deanship of Scientific Research (DSR) at King Abdulaziz University, Jeddah. The authors, therefore, acknowledge with thanks DSR for technical and financial support.
Conflict of interest: None
Financial support: None
Ethics statement: None
A Saadullah, A., & K Abdullah, S. (2014). Detection of Aspergillus species in dried fruits collected from Duhok market and study their aflatoxiginic properties. Rafidain Journal of Science, 25(1), 12-18.
Abdel-Sater, M. A., & Saber, S. M. (1999). Mycoflora and mycotoxins of some Egyptian dried fruits. Bulletin of the Faculty of Sciences, Assiut University, 28(3), 91-107.
Adoum, O. A. (2009). Determination of toxicity levels of some savannah plants using brine shrimp test (BST). Bayero Journal of Pure and Applied Sciences, 2(1), 135-138.
Al Ghamdi, F. L., Bokhari, F. M., & Aly, M. M. (2019). Toxigenic fungi associated with dried Fruits and fruit-based products collected from Jeddah province. Journal Pharm. Biological Sciences, 14(2), 10-20.
Alghalibi, S. M., & Shater, A. R. M. (2004). Mycoflora and mycotoxin contamination of some dried fruits in Yemen Republic. Assiut University Bulletin for Environmental Researches, 7(2), 19-25.
Alhussain, B. S., Alajmi, A. M., Odeh, L. G. H., Nasr, L. E., Awadh, N., & Alotaibi, S. A. A. (2022). Edta vs citric acid decalcifying solutions: a systematic review to compare the clinical efficacy. Annals of Dental Specialty, 10(2), 81-85.
Ali, S., Ali, A., Sartaj, A., Ali, M., & Amjad, A. (2020). Natural occurrence of aflatoxin B1 in dry fruits of Gilgit-Baltistan, Pakistan. Fresenius Environmental Bulletin, 29, 2018-2022.
Al-Jaradi, A., Al-Mahmooli, I., Janke, R., Maharachchikumbura, S., Al-Saady, N., & Al-Sadi, A. M. (2018). Isolation and identification of pathogenic fungi and oomycetes associated with beans and cowpea root diseases in Oman. PeerJ, 6(2), e60-64.
Alp, D., & Bulantekin, Ö. (2021). The microbiological quality of various foods dried by applying different drying methods: a review. European Food Research and Technology, 247, 1333-1343.
Al‐Samarrai, T. H., & Schmid, J. (2000). A simple method for extraction of fungal genomic DNA. Letters in Applied Microbiology, 30(1), 53-56.
Avhad, A., Patil, N., & Zunjarrao, R. (2023). Isolation and Identification of Rhizospheric and Endophytic Fungi Associated with Habenaria brachyphylla (Lindl.) Aitch.: An Endemic and Rare Orchid of the Western Ghats of Maharashtra, India.
Colman, S., Spencer, T. H. I., Ghamba, P. E., & Colman, E. (2012). Isolation and identification of fungal species from dried date palm (Phoenix dactylifera) fruits sold in Maiduguri metropolis. African Journal of Biotechnology, 11(57), 12063-12066.
Costa, A. C. B. P., Rasteiro, V. M. C., Pereira, C. A., Rossoni, R. D., Junqueira, J. C., & Jorge, A. O. C. (2012). The effects of rose bengal‐and erythrosine‐mediated photodynamic therapy on Candida albicans. Mycoses, 55(1), 56-63.
Delcea, C., & Siserman, C. (2020). Validation and Standardization of the Questionnaire for Evaluation of Paraphilic Disorders. Romanian Journal of Legal Medicine, 28(1), 14-20.
Domiaty, D. M. M. (2022). Gum Arabic Mitigates AlCl3-Induced Nephrotoxicity by Upregulating the XRCC1 Gene and Downregulating Ki67 and P53 Expressions. Clinical Cancer Investigation Journal, 11(4), 44-51.
Dsouza, T. S. (2021). Cell Culture and Microscopy as Research Aids in Conservative Dentistry and Endodontics. Annals of Dental Specialty, 9(4), 12-15.
Galea-Holhoș, L. B., Delcea, C., Siserman, C. V., & Ciocan, V. (2023). Age estimation of human remains using the dental system: A review. Annals of Dental Specialty, 11(3), 15.
Gherbawy, Y. A., Elhariry, H. M., & Bahobial, A. A. S. (2012). Mycobiota and mycotoxins (aflatoxins and ochratoxin) associated with some Saudi date palm fruits. Foodborne Pathogens and Disease, 9(6), 561-567.
Gherman, C., Enache, A., & Delcea, C. (2019). The multi-factorial determinism of forensic expertise regarding sentence interruption on medical grounds and decision. Journal of Forensic and Legal Medicine, 61, 45-55.
Govindaraj, A., Paulpandian, S. S., & Shanmugam, R. (2023). Effect of Chlorhexidine and Fluoride Varnish on the Incidence of White Spot Lesion in Orthodontic Patients. Annals of Dental Specialty, 11(1), 35-39.
Hakobyan, L., Grigoryan, K., & Kirakosyan, A. (2010). Contamination of raisin by filamentous fungi–potential producers of ochratoxin A. Potravinárstvo (Slovak Republic), 4(4), 1-8.
Hocking, A. D., Doyle, M. P., Beuchat, M. R., & Montville, T. J. (1997). Toxigenic Aspergillus Species in Food Microbiology-Fundamentals and Frontiers Washington, DC. ASM Press.
Idris, O. A., Wintola, O. A., & Afolayan, A. J. (2019). Evaluation of the bioactivities of Rumex crispus L. leaves and root extracts using toxicity, antimicrobial, and antiparasitic assays. Evidence-Based Complementary and Alternative Medicine, 19(2), 1-10.
Ijabadeniyi, O. A., & Buys, E. M. (2012). Irrigation water and microbiological safety of fresh produce: South Africa as a case study: a review. African Journal of Agricultural Research, 7(35), 4849-4857.
Kallagova, A. R., Bulgakova, Y. V., Oganyan, I. G., Mussakaeva, K. H., Adoneva, V. A., & Burlakova, T. O. (2023). Effectiveness of Fluoridated Milk in Children's Daily Diet for the Prevention of Caries. Archives of Pharmacy Practice, 14(3), 26-32.
Khazamova, S. O., Lazareva, N. V., Semenova, L. V., Lukyanov, G. I., Kolosova, O. Y., & Kamalova, O. N. (2023). Analysis of the Antimicrobial Drugs Market in Russia. Pharmacophore, 14(2), 30-36.
Kumar, R., & Singh, G. (2022). Substituted Benzimidazoles as Antibacterial and Antifungal Agents: A Review. Pharmacophore, 13(2), 41-55.
Lauer, J. (2021). The Response of Salmonella enterica and Enterohemorrhagic Escherichia coli in Low-Moisture Environments (Doctoral dissertation, North Dakota State University).
Momoh, H., Olaleye, A. A., Sadiq, I. S., & Mohammad, A. (2021). Pytochemical Screening, Antimicrobial Activity and Cytotoxicity Effects of Extract of Guiera senegalensis Leaves. ChemSearch Journal, 12(2), 88-93.
Mostafidi, M., Sanjabi, M. R., Shirkhan, F., & Zahedi, M. T. (2020). A review of recent trends in the development of the microbial safety of fruits and vegetables. Trends in Food Science & Technology, 103(2), 321-332.
Najjar, A. A., Alosaimi, E. H., Abduljabbar, H. S., Hashim, H. A., Alem, M. A., Ahmed, M. M. M., & Noor, S. O. (2020). Prevalence of Fungi in Human Follicular Fluid and Its Potential Impact on In Vitro Fertilization Process. Archives of Pharmacy Practice, 11(4), 22-34.
Nicoară, N. D., Marian, P., Petriș, A. O., Delcea, C., & Manole, F. (2023). A review of the role of cognitive-behavioral therapy on anxiety disorders of children and adolescents. Pharmacophore, 14(4), 35-39.
Özer, K. B. (2022). Mycotoxins in Fig. In Advances in Fig Research and Sustainable Production (pp. 318-340). GB: CABI.
Păduraru, L., Jurca, A. D., Zaha, D. C., Ramona, F., Dorobanțu, M. A. H., & Popoviciu, M. S. (2022). Aetiology of Bacterial Infections and Antibiotic Resistance in Pediatric Patients from a Romanian Hospital. Pharmacophore, 13(6), 77-83.
Pavithra, A. S., Paulraj, J., Rajeshkumar, S., & Maiti, S. (2023). Comparative Evaluation of Antimicrobial Activity and Compressive Strength of Conventional and Thyme-Modified Glass Ionomer Cement. Annals of Dental Specialty, 11(1), 70-77.
Radu, C. C., Delcea, C., Plesa, A., & Rad, D. (2023). Transforming perceptions of drug consumption among youth through a Cognitive-Social-Medico-Legal Educational Approach. Pharmacophore, 14(4), 50-56.
Ramadan, E. A., Ramadan, N. A., & Mohammed, A. A. H. (2022). Aflatoxigenic Fungi in Nuts and Dried Fruits in Mosul and Duhok City. Military Medical Science Letters/Vojenské Zdravotnické Listy, 91(3).
Redzuan, A. M., Hui, L. Y., Saffian, S. M., Islahudin, F. H., Bakry, M. M., & Aziz, S. A. A. (2023). Features of Digoxin Toxicity in Atrial Fibrillation and Congestive Heart Failure Patients: A Systematic Review. Archives of Pharmacy Practice, 14(1), 50-55.
Reiss, J. (1972). Vergleichende Untersuchungen über die Toxizität einiger Mykotoxine gegenüber den Larven des Salinenkrebses (Artemia salina L.). Zentralbl Bakteriol Orig, 155(23), 531-534.
Rus, M., Matei, R., Sandu, M. L., Delcea, C., & Siserman, C. (2020). Emotional distress and coping strategies of health care workers during COVID-19 pandemic. Romanian Journal of Legal Medicine, 28, 442-450.
Siserman, C., Delcea, C., Matei, H. V., & Vică, M. L. (2019). Major affective distress in testing forensic paternity. Romanian Journal of Legal Medicine, 27(3), 292-296.
Solis, P. N., Wright, C. W., Anderson, M. M., Gupta, M. P., & Phillipson, J. D. (1993). A microwell cytotoxicity assay using Artemia salina (brine shrimp). Planta Medica, 59(03), 250-252.
Sousa, C. P. D. (2008). The impact of food manufacturing practices on food borne diseases. Brazilian Archives of Biology and Technology, 51(1), 615-623.
Toma, F. M., & Rajab, N. N. (2014). Isolation and identification of fungi from dried fruits and study of quantitative estimation of aflatoxin. Zanco Journal of Pure and Applied Sciences, 26(4), 49-60.
Torres, A. (2007). Food for thought: Microorganism contaminants in dried fruits. California State Science Fair Project Summary. California, 45(3), 1-13.
Tsuneo W. (2010). Pictorial atlas of soil and seed fungi: Morphologies of cultural fungi and key to species. CRC press USA.
Victor, N., Peter, C., Raphael, K., Tendekayi, G. H., Jephris, G., Taole, M., & Portia, P. R. (2017). Microbiological quality of selected dried fruits and vegetables in Maseru, Lesotho. African Journal of Microbiology Research, 11(5), 185-193.
Voiţă-Mekereş, F., Delcea, C., Buhaș, C. L., & Ciocan, V. (2023). Novichok Toxicology: A Review Study. Archives of Pharmacy Practice,14(3), 62-66.
Yuvaraj, A., Priyadharshini, R., Kumar, R., & Sinduja, P. (2023). Anti-Inflammatory and Antifungal Activity of Zinc Oxide Nanoparticle Using Red Sandalwood Extract. Pharmacophore, 14(1), 25-31.
Zhao, Z., Deng, S., Wang, Q., Jia, C., & Yang, J. (2022). Novel Insight into Blocking Cancer Metastasis by Biological Nano Confinement through Altering the Cancer Microenvironment. Clinical Cancer Investigation Journal, 11(4), 10-14.