Computational Repurposing in Computer-Aided Drug Design Against Hantavirus Diseases: A Case Study
|
Venu Paritala* Department of Biotechnology, Vignan's Foundation for Science, Technology & Research (Deemed to be University), Guntur, Andhra Pradesh, India.
|
*E-mail: [email protected]
Abstract
Hantaviruse emerging as a potent zoonotic genus transmitted through rodents, a significant global public health threat, amplifying epidemics with heightened mortality rates. Despite the urgency for effective interventions, the absence of a secure vaccine or antiviral drug against hantavirus disease intensifies the challenge. This study aims to create a strong strategy for computer-aided drug design by utilizing modern computational methodologies. Among the goals include finding critical viral targets, optimizing therapeutic candidates through design, virtual screening of compound libraries, molecular docking simulations, and in silico pharmacokinetics. Computational approaches are chosen for their speed in screening and optimizing new drug candidates, hence speeding up the drug development process. The significance of the study lies in its potential to accelerate the development of treatments for Hantavirus disorders by incorporating computational tools for drug identification and design, offering valuable insights for further experimental validation and clinical outcomes. The goal of this study is to address the critical need for novel techniques to battle the rising global threat of Hantavirus illnesses.
Keywords: Hentavirus, Auto dock, Interactions, Protein-ligand interactions, PymoL
Introduction
Hantaviruses, classified as negative-stranded RNA viruses within the Bunyaviridae family, exhibit distinctive characteristics (Johnson, 2001; Serris et al., 2020). The hantavirus particle is spherical or ovoid, with a diameter of 80–210 nm. Its genome consists of three RNA segments: large (L), medium (M), and small (S). Four structural proteins are expressed by these segments, which together are less than 13 kb in size. The L segment encodes the RNA-dependent RNA polymerase (RdRp), whereas the M segment encodes the glycoproteins Gn and Gc. The S segment encodes the nucleocapsid (N) protein, and the N protein ORF contains a short open reading frame (ORF) that may
represent a putative nonstructural protein (NSs). Critical to the genome's structure are highly conserved consensus sequences at the 3' and 5' ends of all segments, contributing to the formation of a panhandle structure through noncovalent circularization. The genetic material of Hantavirus contains noncoding regions (NCRs) of varying lengths, from 40 to 700 nucleotides (Muyangwa et al., 2015). The longest NCR is found in the S segment, following the N protein ORF. These regions aid in the definition of the various coding sequences in each of the three components of the virus (Alff et al., 2006). Hantavirus replicates in the cytoplasm with the aid of the Golgi system and endoplasmic reticulum (ER). Remarkably, the infected cell appears to be unaffected by the process of Hantavirus replication; in vitro, pathogenic Hantaviruses do not inflict harm or cause programmed cell death (apoptosis) in infected cells. Hantavirus reproduction occurs at a somewhat sluggish rate; in vitro, progeny virus can be found in supernatants from infected cells more than 24 hours after the original infection (Amada et al., 2013). Moreover, the production of hantaviruses from infected cells is still low, which makes it difficult to cultivate high-titer stocks for upcoming studies. It is unclear to what degree this low replication level is unique to in vitro hantavirus growth or if it also occurs in wild hosts and human patients (Baigent et al., 2002), which calls for more research into the complexities of hantavirus replication dynamics.
Hantavirus-Caused Diseases
Hantavirus infections in humans typically occur through the accidental inhalation of rodent manure contaminated with the virus. Human disease is considered incidental, lacking confirmed instances of human-to-natural host transfer (Haller & Kochs, 2002), thereby categorizing humans as dead-end hosts. Human-to-human transmission is infrequent, with rare cases documented for ANDV, and substantial outbreaks remain unrecorded, underscoring a minimal rate of human-to-human infection. However, concerns arise from a recent report indicating PUUV transmission through platelet transfusion, suggesting a potential risk of disease through blood transfusion, akin to other vector-borne viruses.
In Eurasia, where an estimated 150,000 cases emerge annually, the majority manifest as HFRS, predominantly attributed to HTNV in China (Henderson et al., 1995). Up to 10% of patients with severe HFRS caused by DOBV and HTNV may die from hypotensive shock or hemorrhage. The milder clinical course of HFRS brought on by PUUV and SEOV, on the other hand, has a case-fatality rate of between 0.1 and 1%. The Americas report fewer than 1000 instances of HCPS each year. Still, the disease poses a significant health risk, boasting a case fatality rate of up to 40%, with confirmed human-to-human transmission for ANDV.
Renal symptoms characterize HFRS, while HCPS is marked by severe cardiac and pulmonary dysfunction. Although lung and cardiac symptoms may occur in HFRS patients, they are generally less pronounced than in HCPS cases. Renal involvement is also possible in HCPS, albeit less frequently. Both conditions show dynamic phases; HFRS manifests as feverish, hypotonic, oliguric, polyuric, and convalescent phases. But not every site is noticed consistently in every instance. For these conditions, fever, myalgia, headache, and nausea are typical symptoms of the prodromal period. Thrombocytopenia is prevalent in the early stage, coinciding with increased hemorrhagic symptoms. As platelet counts normalize, creatinine levels often rise, signifying renal involvement. The ensuing oliguric phase, characterized by reduced urine output, may necessitate hemodialysis. Subsequent improvement in glomerular filtration marks the polyuric step, featuring increased urine volume. Symptomatic therapy for HFRS varies according to the stage of the illness. PUUV is the main cause of HFRS in Europe, and when bank vole populations rise, the virus spreads among rodents and can be transmitted to humans through epizootics.
Long-Term Consequences of Hantavirus Infection
Traditionally, HFRS and HCPS were perceived as transient illnesses with no enduring consequences for survivors of the acute stage. As such, the focus of medical intervention has been on treating patients with acute HFRS/HCPS to relieve symptoms and eventually lower case fatality rates. Emerging data, however, point to possible long-term effects of hantavirus infections, suggesting that treating acutely infected persons may also help those who are still recovering from HFRS/HCPS. Various publications have explored the potential enduring effects of HFRS, and recent investigations have extended this inquiry to HCPS. Long-term consequences encompass mildly impaired renal and lung function, as well as blood pressure fluctuations, with some cases requiring hormone replacement therapy for chronic hormonal deficits (Schönrich et al., 2008). A 10-year follow-up study following PUUV infection revealed no discernible lasting repercussions, challenging the notion that hantaviruses invariably lead to long-term complications. Furthermore, new research shows no association between the severity of acute PUUV-induced HFRS and the patients' long-term results. This implies that, if hantavirus infections do have long-term effects, the long-term prognosis is influenced by unknown variables. Interestingly, viremia during HFRS/HCPS is thought to be temporary, in contrast to what has been observed in natural hosts.
Vaccines/Therapeutics Currently Available
To be useful, countermeasures against hantavirus infection should be able to prevent exposure, limit the development of a subsequent disease in people after susceptibility, or lessen the likelihood of a fatal result while mitigating symptoms. As of right now, there are no FDA-approved drugs available in the US to treat HFRS or HCPS (Hepojoki et al., 2012; Hepojoki et al., 2014). Patients are mostly treated for symptoms; at this time, there are no known medicines that target the replication or hantavirus proteins, with the possible exception of ribavirin, a nucleoside analog, for HFRS. Vaccinating susceptible populations may seem like a good preventative measure.
Numerous vaccination methods have been used in experiments, and numerous vaccine candidates have been investigated in human subjects. Though similar immunizations are also available in China, Hantavax, a suckling mouse brain-derived vaccine, is only available in Korea. As of right now, there isn't much evidence to support the vaccines' ability to prevent HFRS. With varying degrees of efficacy, ribavirin has been used to treat HFRS and HCPS. In clinical studies, ribavirin is effective against HFRS but not HCPS. It has not been utilized to treat patients with ANDV, DOBV, PUUV, or SEOV infections. Except for ribavirin, several antiviral treatments have been investigated in vitro and in animal models but have not been evaluated in humans.
Hantavirus Immunology: HFRS/HCPS Findings
Hantaviruses cannot spread from natural hosts to humans unless they are breathed in, cause a systemic infection, and then are expelled into the saliva, urine, or feces. The virus must first overcome early immunological barriers such as interferons (IFNs) produced in infected cells, antiviral peptides/proteins in body fluids, and apoptosis induced by infected cells or natural killer cells. Effective infection triggers antigen-specific (adaptive) immune responses that the virus has to fend off. Both the innate and adaptive immune systems are strongly engaged during acute infection. Hantaviruses can decrease immune responses by actively interfering with cellular signaling pathways and other cellular functions. It appears that hantaviruses have developed a resistance to the antiviral effects of saliva and maybe other physiological fluids since hantaviruses are generally insensitive to saliva and to certain human salivary components known to have antiviral action against other viruses. The only hantavirus that is known to spread between humans is ANDV. ANDV infection may run in families, and sexual partners of those with HCPS have the highest chance of contracting the virus, indicating that close contact with infected individuals is necessary for ANDV human-to-human transmission.
ANDV-infected HCPS patients excrete the infectious virus in their urine, yet it is still unknown how ANDV is transmitted from person to person. The preferred mode of transmission for ANDV amongst its native host, long-tailed rice rats, maybe oral. Up to nine days after the onset of the disease, we detected PUUV RNA in the saliva of HFRS patients. Nevertheless, when injecting bank voles with HFRS patients' saliva, no infectious virus was found, indicating that the PUUV RNA found in saliva may have been non-infectious viral particles. While it has not been investigated whether human saliva spreads ANDV, it is more resistant than PUUV and HTNV to the antiviral effects of saliva. IFNs have a major role in inducing antiviral responses in both bystander and infected cells. After attaching to viral components, cellular pattern recognition receptors (PRRs) trigger signaling pathways that result in the production of type I and III IFNs (IFN-a/b/l). The primary PRRs for negative-stranded RNA viruses are TLR3, RIG-I, and MDA-5. The PRRs that recognize hantaviruses are still unknown, and HFRS/HCPS-causing hantaviruses are known to be poor type I IFN (IFN-a/b) inducers in vitro. IFN-a, -b, and -l levels are not increased in PUUV-infected individuals.
Literature Review
The organization of viral particles, in particular the role of the hantavirus membrane fusion glycoprotein Gc, which is essential for virus entry, is poorly understood. The type of hantavirus that causes HFRS, the Hantaan virus, has Gc in neutral pH, monomeric pre-fusion, acidic pH, and trimeric post-fusion forms. We present the X-ray structures of these forms for your reference (Jonsson et al., 2008). With the characteristic --sheet domains designated I, II, and III initially discovered in arbovirus fusion proteins like alpha- and flaviviruses, the structures demonstrate that Gc is a class II fusion protein. The structures also exhibit certain Gc-specific features that set them apart from the proteins of the arbovirus class II. By inserting residues from three distinct loops into the target membrane, hantavirus Gc causes fusion rather than a single "fusion loop," as demonstrated functionally by structure-guided mutagenesis on the Andes virus that induces HPS. We further demonstrate that only at an acidic pH does the membrane-contacting area of Gc become structured via a mix of polar and electrostatic interactions.
The laboratory findings included leukocytosis (Gavrilovskaya et al., 1998; Bi et al., 2008; Heyman et al., 2009; Chen et al., 2011), often with myeloid precursors, an increased hematocrit, thrombocytopenia (median lowest platelet count, 64,000 per cubic millimeter), prolonged prothrombin and partial thromboplastin times, an elevated serum lactate dehydrogenase concentration, decreased serum protein concentrations, and proteinuria (Kale et al., 2022). 15 of the 17 patients (88 percent) suffered rapidly increasing acute pulmonary edema, and 13 died, all of whom had significant hypotension (case fatality rate, 76 percent). Increases in hematocrit and partial thromboplastin time were predictive of mortality. In addition, the structure shows that, in comparison to traditional class II fusion proteins, hantavirus Gc has an additional N-terminal "tail" that is essential for stabilizing the post-fusion trimer as well as a relocation of domain III in the trimer's quaternary organization. It is predicted that the mechanistic insights gleaned from this data will provide a clear understanding of how to create treatments for these illnesses in humans. The patients were 61% female, 72% Native American, 22% Caucasian, and 6% Hispanic, with an average age of 32.2 years (range: 13 to 64). Fever and myalgia (100%), cough or dyspnea (76%), gastrointestinal symptoms (76%), and headache (76%), were the most common prodromal symptoms. (71 percent). Among the physical indicators, tachypnea (100%), tachycardia (94%), and hypotension were the most common (50 percent).
Materials and Methods
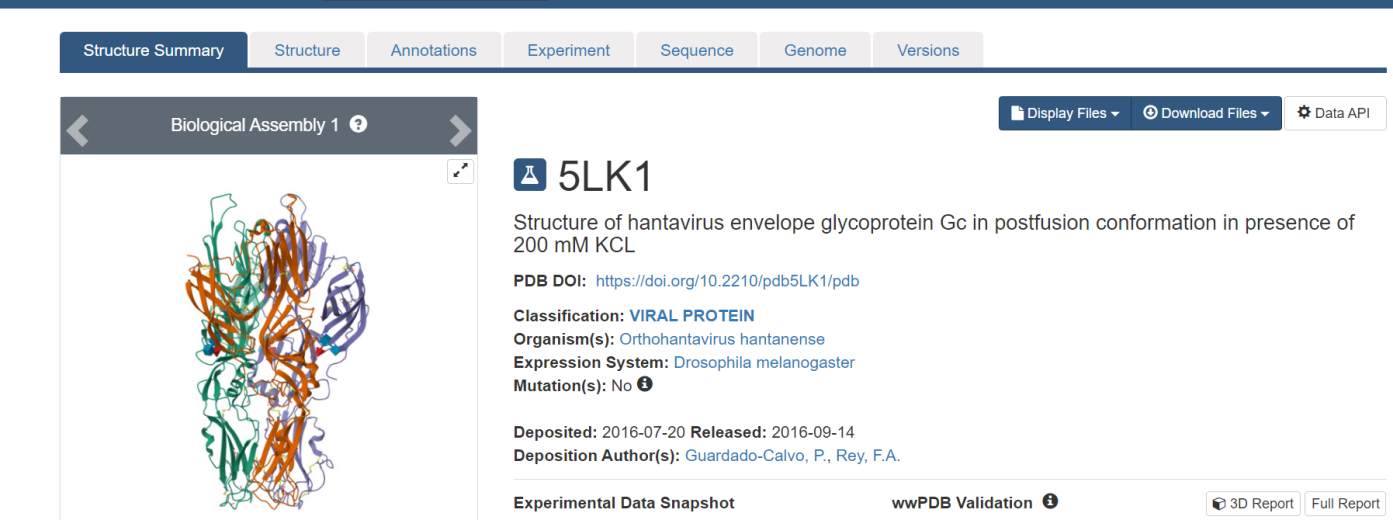
The three-dimensional structure data for important biological elements, such as proteins and nucleic acids, may be found in the Protein Data Bank (PDB), where the Hantavirus structural data was obtained (Paritala et al., 2022). This priceless database gathers information from a variety of techniques, including NMR spectroscopy, cryo-electron microscopy, and X-ray crystallography. Biologists and biochemists worldwide contribute to this repository, submitting their findings. Accessible freely on the Internet through member organization websites (PDBe, PDBj, RCSB, and BMRB), the PDB offers a comprehensive resource for researchers. To locate our specific protein of interest, we searched within the PDB and downloaded the structural data in PDB format, as illustrated in Figure 1.
|
|
|
Figure 1. 5LK1 strature of hantavirus envelope Glycoprotein Gc |
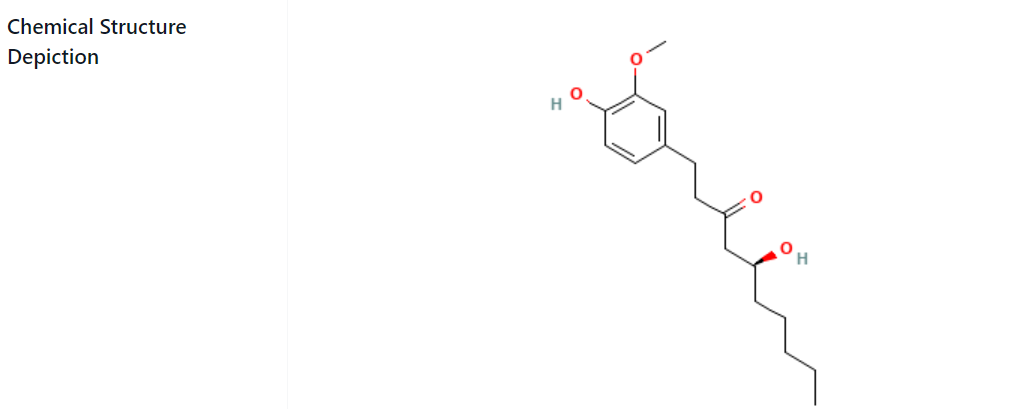
Gingerol
Since ancient times, humans have utilized Ginger, scientifically known as Zingiber officinale, not only for culinary purposes to enhance the taste of dishes but also for its medicinal properties. Ginger encompasses various phenolic compounds, including gingerol, shogaol, and paradol, each exhibiting antioxidant, anti-tumor, and anti-inflammatory activities. Over the past two decades, numerous experts have delved into the study of Ginger and its components, providing substantial scientific evidence supporting its health benefits (Siserman et al., 2019; Galea-Holhoș et al., 2023; Voiţă-Mekereş et al., 2023). This review focuses on the research findings and endeavors dedicated to gingerols, the primary aromatic constituents of Ginger. It explores how gingerols modify and target signaling pathways, inducing changes that alleviate, reverse, or prevent chronic illnesses in both human studies and animal models (Nicoară et al., 2023). The physical, chemical, and biological characteristics of gingerols are thoroughly examined. The utilization of Ginger, particularly gingerols, as a medical food derivative, is a safe approach to the treatment or prevention of chronic illnesses, promising benefits for the general public, medical professionals, patients, researchers, students, and industrialists. The traget molecule of Gingerol was retrived from Pubchem (Figure 3).
Autodock Vina
Autodock, a specialized simulation software designed for molecular docking, significantly focuses on facilitating interactions between proteins and ligands. Renowned for its user-friendly interface and open-access availability, Autodock stands as a well-established and widely utilized docking program. Successful molecular docking simulations using Autodock necessitate the possession of both the software for preparing and evaluating AutoDock dockings along with the Autodock program itself (Gherman et al., 2019; Rus et al., 2020). The comprehensive AutoDock package encompasses both the AutoDock and AutoGrid applications. While AutoDock manages the docking process, it relies on grid maps generated by AutoGrid to define the field of interaction energies surrounding the macromolecular target of interest (Figure 2). This process expedites the computation of interaction energy. The pre-computed maps, skillfully crafted by AutoGrid, offer versatility and can be applied across an unlimited number of dockings. Additionally, a dedicated graphical user interface (GUI) named AutoDockTools is provided separately, enhancing user interaction and control.
|
|
|
Figure 2. Protein vandalizing |
Pymol
PyMOL stands as a free and open-source molecular visualization system, proficient in generating high-quality 3D images encompassing small compounds and biological macromolecules such as proteins. Remarkably, the original author notes that, by 2009, PyMOL played a pivotal role in crafting nearly a quarter of all published 3D protein structure images in the scientific literature (Delcea & Siserman, 2020; Radu et al., 2023). In the realm of structural biology, PyMOL stands out as one of the few selected open-source tools for model visualization. The "Py" in PyMOL is a nod to its creation in the Python programming language. Leveraging the OpenGL Extension Wrangler Library (GLEW), FreeGLUT, and the Adaptive Poisson Boltzmann Solver for solving Poisson-Boltzmann equations, PyMOL demonstrates its versatility. An intriguing application involves utilizing docking files extracted from AutoDock Vina, subsequently employing PyMOL for the visualization of the model, showcasing the software's capability in structural analysis.
|
|
|
Figure 3. Target Molecule |
Results and Discussion
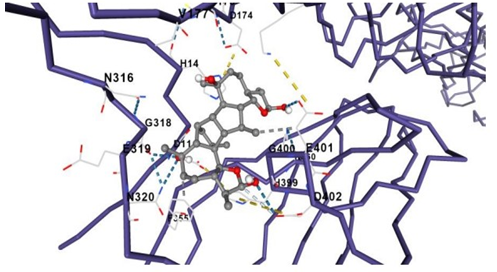
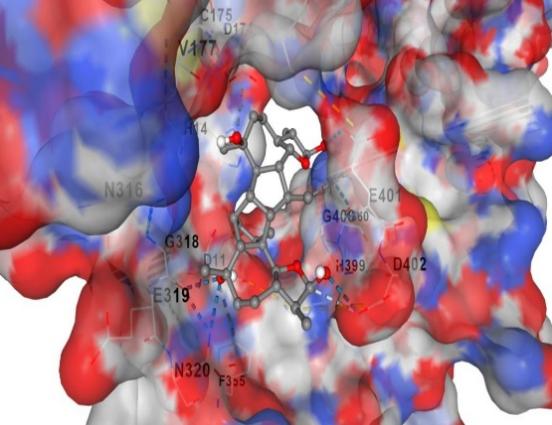
The dlg log file contains docking results. This diagram depicts the configuration of the ligand attached to 5Lk1 from 1 to 15. Concerning the reference structure, the optimal conformation has a binding energy (G) of -8.4 kcal/mol and an inhibitory constant cavity size of 139 (Table 1). This proves the accuracy and dependability of Autodock's results. New 5Lk1 inhibitors could be found with the use of docking and virtual screening (Figure 4).
|
|
|
a) |
|
|
|
b) |
|
Figure 4. a, b) Binding site of Taget Protein |
Table 1. Binding energies of Ligands docked agains Hantavirus
|
11 Vina |
11 Cavity |
Center |
|
Size |
|||
|
score |
size |
X |
y |
|
x |
y |
Z |
|
-8.4 |
139 |
-39 |
103 |
32 |
22 |
22 |
22 |
|
-8.4 |
83 |
-34 |
101 |
29 |
22 |
22 |
22 |
|
-7.4 |
114 |
-49 |
100 |
19 |
22 |
22 |
22 |
|
-7.2 |
822 |
-44 |
107 |
20 |
22 |
22 |
22 |
|
-7.1 |
71 |
-42 |
110 |
42 |
22 |
22 |
22 |
|
-6.9 |
189 |
-60 |
103 |
6 |
22 |
22 |
22 |
|
-6.9 |
58 |
-53 |
117 |
3 |
22 |
22 |
22 |
|
-6.7 |
1493 |
-42 |
107 |
-2 |
22 |
22 |
28 |
|
-6.6 |
75 |
-46 |
116 |
-9 |
22 |
22 |
22 |
|
-6.5 |
351 |
-38 |
114 |
15 |
22 |
22 |
22 |
|
-6.4 |
63 |
-50 |
115 |
-22 |
22 |
22 |
22 |
|
-6.2 |
67 |
-55 |
107 |
51 |
22 |
22 |
22 |
|
-6.1 |
105 |
-34 |
111 |
24 |
22 |
22 |
22 |
|
-6 |
103 |
-53 |
104 |
-51 |
22 |
22 |
22 |
|
-5.7 |
70 |
-44 |
97 |
51 |
22 |
22 |
22 |
In recent years, the HantaVirus strain has been seen as a threat to disease control and has piqued the interest of national and international health agencies. Numerous mutational events have been found, which may result in resistance to a medicine routinely used to treat Hanta. These stories underscore the importance of further investigation into alternative therapy modalities.
This effort aimed to use the AutoDock Vina tool to provide the groundwork for in vitro studies from the perspective of protein-ligand molecular computing. The results produced by AutoDock Vina software are of the same caliber as those produced by AutoDock software, notwithstanding the speed at which calculations are completed. Another goal was to dispel the myth about the possible health benefits of flavonoid ingestion on the Hantavirus.
Conclusion
We conducted docking experiments with hantaviral protein and antiviral plant compounds (6-Gingerol). Ginger includes a variety of phenolic chemicals, including gingerol, shogaol, and paradol, which have antioxidant, anti-tumor, and anti-inflammatory activities. Using the symbol and auto dock utilities, the findings indicate that the (high affinity) binding zone is located distant from the active site. This diagram depicts the configuration of the ligand attached to 5Lk1 from 1 to 15. The optimum conformation has an inhibition constant cavity size 139 from the reference structure of 1.22 and a binding energy (G) of 8.4 kcal/mol. Several occurrences of mutations have been recorded, which may result in resistance to a medicine routinely used to treat Hantavirus.
Acknowledgments: The authors would like to thank Vignan's Foundation for Science, Technology & Research (Deemed to be University) for giving Technical support.
Conflict of interest: None
Financial support: None
Ethics statement: None
Alff, P. J., Gavrilovskaya, I. N., Gorbunova, E., Endriss, K., Chong, Y., Geimonen, E., Sen, N., Reich, N. C., & Mackow, E. R. (2006). The pathogenic NY-1 hantavirus G1 cytoplasmic tail inhibits RIG-I-and TBK-1-directed interferon responses. Journal of Virology, 80(19), 9676-9686. doi:10.1128/JVI.00508-506
Amada, T., Yoshimatsu, K., Yasuda, S. P., Shimizu, K., Koma, T., Hayashimoto, N., Gamage, C. D., Nishio, S., Takakura, A., & Arikawa, J. (2013). Rapid, whole blood diagnostic test for detecting anti-hantavirus antibody in rats. Journal of Virological Methods, 193(1), 42-49. doi:10.1016/j.jviromet.2013.04.02
Baigent, S. J., Zhang, G., Fray, M. D., Flick-Smith, H., Goodbourn, S., & McCauley, J. W. (2002). Inhibition of beta interferon transcription by noncytopathogenic bovine viral diarrhea virus is through an interferon regulatory factor 3-dependent mechanism. Journal of Virology, 76(18), 8979-8988. doi:10.1128/JVI.76.18.8979-8988.2002
Bi, Z., Formenty, P. B., & Roth, C. E. (2008). Hantavirus infection: a review and global update. The Journal of Infection in Developing Countries, 2(01), 003-023.
Chen, C. C., Pei, K. C., Yang, C. M., Kuo, M. D., Wong, S. T., Kuo, S. C., & Lin, F. G. (2011). A possible case of hantavirus infection in a Borneo orangutan and its conservation implication. Journal of Medical Primatology, 40(1), 2-5. doi:10.1111/j.1600-0684.2010.00442.x (2010)
Delcea, C., & Siserman, C. (2020). Validation and Standardization of the Questionnaire for Evaluation of Paraphilic Disorders. Romanian Journal of Legal Medicine, 28(1), 14-20.
Galea-Holhoș, L. B., Delcea, C., Siserman, C. V., & Ciocan, V. (2023). Age estimation of human remains using the dental system: a review. Annals of Dental Specialty, 11(3), 14-18.
Gavrilovskaya, I. N., Shepley, M., Shaw, R., Ginsberg, M. H., & Mackow, E. R. (1998). β3 integrins mediate the cellular entry of hantaviruses that cause respiratory failure. Proceedings of the National Academy of Sciences, 95(12), 7074-7079. doi:10.1073/pnas.95.12.7074
Gherman, C., Enache, A., & Delcea, C. (2019). The multi-factorial determinism of forensic expertise regarding sentence interruption on medical grounds and decision. Journal of Forensic and Legal Medicine, 61, 45-55.
Haller, O., & Kochs, G. (2002). Interferon‐induced mx proteins: dynamin‐like GTPases with antiviral activity. Traffic, 3(10), 710-717. doi:10.1034/j.1600- 0854.2002.31003
Henderson, W. W., Monroe, M. C., Jeor, S. C. S., Thayer, W. P., Rowe, J. E., Peters, C. J., & Nichol, S. T. (1995). Naturally occurring Sin Nombre virus genetic reassortants. Virology, 214(2), 602-610. doi:10.1006/viro.1995.0071
Hepojoki, J., Strandin, T., Lankinen, H., & Vaheri, A. (2012). Hantavirus structure–molecular interactions behind the scene. Journal of General Virology, 93(8), 1631-1644. doi:10.1099/vir.0.042218-10
Hepojoki, J., Vaheri, A., & Strandin, T. (2014). The fundamental role of endothelial cells in hantavirus pathogenesis. Frontiers in Microbiology, 5, 727. doi:10.3389/fmicb.2014.0072
Heyman, P., Vaheri, A., Lundkvist, Å., & Avsic-Zupanc, T. (2009). Hantavirus infections in Europe: from virus carriers to a major public-health problem. Expert Review of Anti-Infective Therapy, 7(2), 205-217.
Johnson, K. M. (2001). Hantaviruses: history and overview. Hantaviruses, 256, 1-14.
Jonsson, C. B., Hooper, J., & Mertz, G. (2008). Treatment of hantavirus pulmonary syndrome. Antiviral Research, 78(1), 162-169.
Kale, B. S., Bhale, M. S., Bhagat, A. B., & Khairnar, S. A. (2022). Pharmacognostic evaluation of osyris quadripartita salz. ex decne. Pharmacophore, 13(3), 50-56.
Muyangwa, M., Martynova, E. V., Khaiboullina, S. F., Morzunov, S. P., & Rizvanov, A. A. (2015). Hantaviral proteins: structure, functions, and role in hantavirus infection. Frontiers in Microbiology, 6, 1326. doi:10.3389/fmicb.2015.0132
Nicoară, N. D., Marian, P., Petriș, A. O., Delcea, C., & Manole, F. (2023). A review of the role of cognitive-behavioral therapy on anxiety disorders of children and adolescents. Pharmacophore, 14(4), 35-39.
Paritala, V., Thummala, H., & Mohith, T. N. S. (2022). Homology Modeling and Molecular Docking Studies of Selected Substituted Tetradecane on vlsE Borrelia spielmanii. Journal of Molecular Docking, 2(1), 16-28.
Radu, C. C., Delcea, C., Plesa, A., & Rad, D. (2023). Transforming perceptions of drug consumption among youth through a Cognitive-Social-Medico-Legal Educational Approach. Pharmacophore, 14(4), 50-6.
Rus, M., Matei, R., Sandu, M. L., Delcea, C., & Siserman, C. (2020). Emotional distress and coping strategies of health care workers during COVID-19 pandemic. Romanian Journal of Legal Medicine, 28, 442-450.
Schönrich, G., Rang, A., Lütteke, N., Raftery, M. J., Charbonnel, N., & Ulrich, R. G. (2008). Hantavirus‐induced immunity in rodent reservoirs and humans. Immunological Reviews, 225(1), 163-189.
Serris, A., Stass, R., Bignon, E. A., Muena, N. A., Manuguerra, J. C., Jangra, R. K., Li, S., Chandran, K., Tischler, N. D., Huiskonen, J. T., et al. (2020). The hantavirus surface glycoprotein lattice and its fusion control mechanism. Cell, 183(2), 442-456. doi:10.1016/j.cell.2020.08.023
Siserman, C., Delcea, C., Matei, H. V., & Vică, M. L. (2019). Major affective distress in testing forensic paternity. Romanian Journal of Legal Medicine, 27(3), 292-296.
Voiţă-Mekereş, F., Delcea, C., Buhaș, C. L., & Ciocan, V. (2023). Novichok Toxicology: A Review Study. Archives of Pharmacy Practice, 14(3), 62-66.