Chemical Components of Polygonum cuspidatum Ethylacetate Subfraction and their Effects on Epstein-Barr Virus Lytic Genes Expression
Ching-Yi Yiu, Yi-Hsuan Kuan, Yi-Jie Chen, Bo-Shine Wu, Tsuey-Pin Lin*
Abstract
The study highlights the effect of ethylacetate subfraction F1 from Polygonum cuspidatum root and piceid on the inhibition of Epstein-Barr virus (EBV) lytic gene expression. The MTT [3-(4,5-dimethyldiazol-2-yl)-2,5-diphenyltetrazolium bromide] method was employed to assess the effect of F1 and piceid on the cell survival of P3HR1. The immunoblotting method and flow cytometry were applied to analyze the expression of EBV lytic proteins. The results showed that the EC50 (effective concentration required to inhibit 50% EBV lytic proteins) of F1 and piceid inhibiting the lytic proteins of EBV were 23.3 μg/mL and 119.6 μM, respectively. For P3HR1 cytotoxicity, the CC50 (cytotoxic concentration that decreased cell viability by 50%) of F1 and piceid were 93.1 μg/mL and 507.9 μM, respectively. In other words, F1 and piceid effectively inhibited the expression of EBV lytic proteins at a non-cytotoxic concentration. qPCR was performed to analyze the effect of F1 and piceid on the transcription of lytic genes, including BRLF1, BZLF1, and DNA replication of EBV. The results showed that F1 and piceid inhibited the transcription of lytic genes and reduced DNA replication of EBV with EC50 of 55.5 μg/mL and 89.9 μM, respectively. The results of this study confirmed that F1 and piceid could effectively inhibit the expression of EBV lytic proteins and EBV DNA replication, indicating that F1 and piceid are potentials for use as anti-EBV drugs.
Keywords: Piceid, Antiviral activity, Epstein-Barr virus, Polygonum cuspidatum
Introduction
Epstein-Barr virus (EBV) is a human herpesvirus that belongs to the g-herpes virus subfamily. EBV mainly infects human B lymphocytes and epithelial cells causing infectious mononucleosis (Murata et al., 2021) and many malignant diseases such as Burkitt's lymphoma (Heslop, 2020), Hodgkin's disease, gastric cancer, T-cell lymphomas and nasopharyngeal carcinoma (Wong et al., 2022). When cells are infected with EBV, virion production is reduced during the latent cycle, presenting the coexistence of virus and host (Murata et al., 2021). The EBV is reactivated by ultraviolet light (Chen et al., 2020), sodium butyrate (SB) (Koike et al., 2020), 12-O-tetradecanoyl phorbol 13-acetate (TPA) (Dochi et al., 2022), trichostatin A (TSA), and valproic acid (VPA) (Anderson et al., 2019) allowing the BZLF1 and BRLF1 genes to express immediate-early proteins Zta and Rta, respectively. Zta and Rta are transcription factors that can activate the transcription of BMRF1 and BALF5, which encode the diffused early antigen (EA-D) and DNA polymerase and promote EBV’s entering the lytic cycle (Murata et al., 2021). The viral DNA replicates by a rolling circle mechanism and assembles into virus capsid antigens and membrane proteins to produce mature virus particles; EB virions then infect other B lymphocytes and epithelial cells.
Some studies have found that EBV entering the lytic cycle can induce cytokines and cause tumorigenesis. For example, the EBV immediate early lytic protein Zta induces the production of multiple oncogenic and inflammatory factors (Jangra et al., 2019). Therefore, inhibiting the virus entry into the lytic cycle can effectively treat EBV-related diseases. Nucleoside analogs, such as acyclovir and ganciclovir, are often used to treat EBV infection and are effective in treating infectious mononucleosis (Zhang et al., 2021), post-transplant lymphoproliferative disorder (Dugan et al., 2018). Earlier studies have shown that acyclovir and ganciclovir can affect EBV replication by inhibiting virus-encoded DNA polymerase (Pagano et al., 2018) but not the expression of EBV's immediate early and early lytic proteins. In addition to chemical substances, ingredients extracted from plants can inhibit the lytic cycle, such as epigallocatechin gallate (EGCG), resveratrol, curcumin, sulforaphane, protoapigenone, luteolin, andrographolide, moronic acid, emodin, and glycyrrhizic acid (Yiu et al., 2010; Yiu et al., 2014; Andrei et al., 2019; Wu et al., 2019; Benedetti et al., 2020; Li et al., 2021; Shao et al., 2022). They prevent EBV from entering the lytic cycle by blocking the expression of EBV early proteins.
Earlier studies exhibit that the ethanolic extract of P. cuspidatum and resveratrol inhibit EBV lytic cycle and induce EBV-positive cell apoptosis (Yiu et al., 2010; Yiu et al., 2014; Wu et al., 2018; Yiu et al., 2021). The ethanolic extract of P. cuspidatum root contains anthraglycoside B, physcion, piceid, emodin, and resveratrol (Yun-Ting et al., 2020). Piceid (3,5,4-trihydroxystilbene-3-O-β-D-glucopyranoside) also called polydatin, is a resveratrol derivative, which retains the biological activity of resveratrol; it has the functions of anti-inflammation, regulating the immune system, antioxidation, inducing tumor cell apoptosis, inhibiting genetic mutations, preventing cardiovascular disease, protecting myocardial and liver cells, lowering cholesterol in the blood, inhibiting lipid peroxidation, and preventing shock (Karami et al., 2022). It can be found in plants such as peanuts, Polygonum cuspidatum, mulberry, grape skin, and cocoa beans. To date, no studies have been conducted on the inhibition of immediate-early gene expression and DNA replication of EBV by piceid. This study will explore whether P. cuspidatum root subfraction F1 and piceid can affect the DNA replication of EBV by blocking early gene transcription of EBV and expression of lytic proteins (Aiche et al., 2022; Almaghrabi et al., 2022; Chandra & Meel, 2022; Fayiah et al., 2022; Pimple, 2022).
Materials and Methods
Plant Material
P. cuspidatum was collected from the San-Dei-Men area in Pingtung County in Taiwan and verified by Prof. C. S. Kuoh. The specimen was deposited in the herbarium of the National Cheng Kung University, Tainan, Taiwan. Piceid was purchased from Sigma Chemical Co. (St. Louis, Mo, USA).
Sample Preparation and HPLC Analysis
Using the Yiu method (Yiu et al., 2014), the subfraction F1 of ethyl acetate isolated from P. cuspidatum root was prepared and analyzed by high-performance liquid chromatography with a diode array detector ranging from 210 to 500 nm. A yield of 1.81% was obtained. The resulting residues were finally dissolved in dimethyl sulfoxide (DMSO).
Cell Culture and Lytic Induction of EBV
P3HR1, a Burkitt’s lymphoma cell line latently infected by EBV, was cultured in RPMI 1640 medium containing 10% fetal calf serum (Biological Industries, Israel). Cells were treated with 3 mM of sodium butyrate (SB) to induce the EBV lytic cycle (Yiu et al., 2010).
Cytotoxicity Assay
P3HR1 cells were treated with Polygonum cuspidatum root subfraction F1 or piceid. After 24 h incubation, a 0.5 mg/mL [3-(4,5-dimethyldiazol-2-yl)-2,5 diphenyl tetrazolium bromide (MTT) solution was added into RPMI 1640 medium. The dehydrogenase activity representing the viable cells was measured.
Immunoblot Analysis
The cell lysate was prepared from 3 x 106 P3HR1 cells with 100 μl of lysis buffer that contained 50 mM Tris-HCl, pH 7.8, 150 mM NaCl, 5 mM EDTA, 0.5% Triton X-100 and 0.5% NP40 using a method described elsewhere (Yiu et al., 2010). SDS-polyacrylamide gel electrophoresis and immunoblot analysis with anti-Rta, anti-Zta, and anti-EA-D antibodies purchased from Argene (Varilhes, France) were performed using methods described previously.
RNA and DNA Extraction
For qPCR, RNA was extracted from 3 × 106 cells using the conventional Trizol method (Invitrogen, Carlsbad, CA). DNA removal was performed using the RNase-Free DNase Set (promega, Madison, WI). Total RNA was added to the High capacity cDNA reverse transcription kit (Applied Biosystems) to produce cDNA. Briefly, 10 μL of treated RNA (2 μg) was mixed with 0.8 μL 25 × dNTP mix (100 mM) and 2 μL 10 × RT Random Primers, followed by adding 2 mL 10 × RT buffer, 4.2 μl 0.1% DEPC water and 1 μL MultiScribe™ Reverse Transcriptase (50 U/μL). The components in the tube were mixed gently and incubated for 10 minutes at 25℃, for 120 minutes at 37℃, and 5 minutes at 85℃. DNA was extracted using a Quick-gDNA miniPrep kit (Zymo Research).
Real-Time Quantitative PCR
For mRNA analysis, Quantitative PCR (qPCR) was done with the SYBR green (Applied Biosystems); DNA analysis using Taqman probe was performed (5′-6-FAM-GGAGACACATCTGGACCAG-MGBNFQ-3′) on the ABI StepOneTM qPCR system with StepOneTM software v 2.1. All reactions were run in triplicate. Mean cycle threshold (CT) values were normalized to β-actin, yielding a normalized CT (ΔCT). The ΔΔCT value was calculated by subtracting respective control from the ΔCT, and the expression level was then calculated by 2 raised to the power of the respective −ΔΔCT value. Relative mRNA (DNA) level (%) = 2-ΔΔCT (SB and sample treatment)/ 2-ΔΔCT (SB treatment) × 100. Primers for mRNA analysis include the following: BRLF1 forward (5′-TCACTACACAAACAGACGCAGCCA-3′) and reverse (5′-AATCTCCACACTCCCGGCTGTAA-3′); BZLF1 forward (5′-AGAAGCACCTCAACCTGGAGACAA-3′) and reverse (5′-CAGCGATTCTGGCTGTTGTGGTTT-3′); and β-actin forward (5′-CGTCTTCCCCTCCATCG) and reverse (5′-CTCGTTAATGTCACGCAC-3′). Primers for DNA analysis, EBNA1 forward (5′-TACAAGACCTGGAAAGGCC-3′) and reverse (5′-TCTTTGAGGTCCACTGCC -3′).
Statistical Analysis
One-way analysis of variance (ANOVA) was used to analyze the data followed by Dunnett’s post hoc test using SAS JMP 8.0 software. Values are expressed as mean ± SD of three replicates and a p-value of <0.05 is regarded as significant.
Results and Discussion
Composition Analysis of Polygonum cuspidatum Ethylacetate Subfraction F1
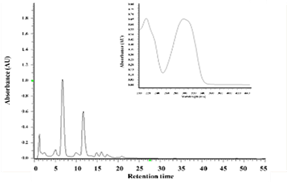
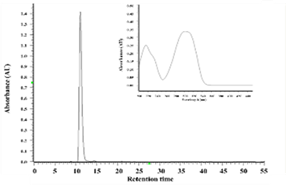
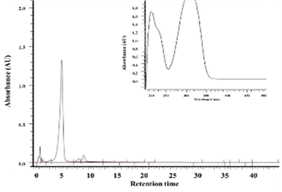
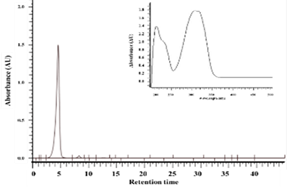
The ethanol extract of Polygonum cuspidatum root was subject to partition into hexane and ethyl acetate, and the collected ethyl acetate fraction was separated by a semi-preparative high-pressure liquid chromatography analyzer to obtain the ethyl acetate subfraction F1 and F1a. Reverse-phase high-performance liquid chromatography was used to analyze the composition of F1 and F1a; values obtained were compared with the retention time and ultraviolet spectrum of pure standards of piceid and resveratrol to determine the target compounds in F1 and F1a. As shown in Figure 1, the peak eluting at 5.13 min was piceid and at 11.6 min was resveratrol. The content of piceid and resveratrol in F1 was 20.49 and 10.4% and in F1a were 51.76% and 1.17%, respectively.
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
Figure 1. HPLC chromatograms and UV spectra of the ethylacetate subfraction F1 (a) and F1a (c) isolated from Polygonum cuspidatum root (a) and pure standard of resveratrol (b) and piceid (d) |
Effect of F1 and Piceid on the Growth of P3HR1 Cells
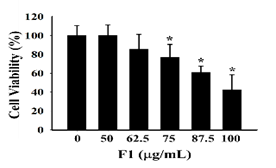
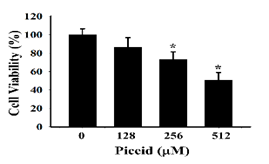
To understand whether the ethyl acetate subfraction F1 from the roots of Polygonum cuspidatum and piceid have antiviral activity without affecting cell survival, the MTT method was proposed to analyze the survival rate of P3HR1 cells. After 24 hours of adaptation, P3HR1 cells (1 × 105 cells/mL) were added to different concentrations of F1 (0-100 μg/mL) or piceid (0-512 μM); after 24 hours of reaction, the MTT method was then performed to determine the cell survival rate. The results showed that F1 and piceid did not affect the survival rate of P3HR-1 cells. When concentrations of F1 and piceid were below 62.5 μg/mL and 128 μM, respectively, the cell survival rate was higher than 85%. When the F1 concentration was increased to 100 μg/ml, the cell survival rate was only 40%. Piceid affected the survival rate of P3HR1 cells at the concentration of 256 μM and 512 μM, showing 72.9% and 50.4%, respectively. The median cytotoxic concentration (CC50) of F1 and piceid were 93.1 μg/mL and 507.9 μM, respectively, which is shown in Figure 2.
|
|
|
a) |
|
|
|
b) |
|
Figure 2. Toxicity of F1 and piceid on P3HR1 cells |
Use of Immunoblotting to Assess the Effect of F1 and Piceid on EBV Lytic Proteins Expression
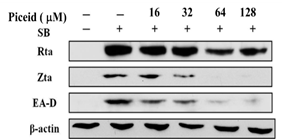
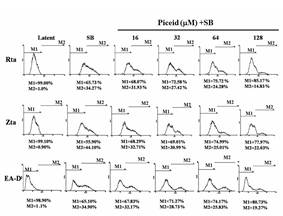
EBV reactivation can be induced by sodium butyrate to express the immediate-early proteins, Zta and Rta, which are transcription factors involved in the promotion of the expression of other lytic cycle genes, such as activation of the translation of the BRLF1 gene to produce EA-D protein. EBV is reactivated from the latent to lytic cycle and infects other cells. Therefore, EBV lytic protein expression represents that EBV is in the DNA replication stage. 6 × 105/mL of P3HR1 cells were treated with the ethylacetate subfraction F1 from Polygonum cuspidatum root and piceid. Before the induction of the EBV lytic cycle, SB was added. After 24-hour treatment, the cells were harvested. The lytic proteins, Rta, Zta, and EA-D of EBV in the cells were determined by immunoblot assay. Figure 3 indicates that F1 and piceid significantly inhibited the expression of Rta, Zta, and EA-D at the concentration of 12.5 μg/mL and 32 μM, respectively. The F1 concentration increasing to 25 μg/mL and 64 μM for piceid completely inhibited the expression of Zta and EA-D. The performance of Rta, Zta, and EA-D is no longer detectable at 50 μg/mL for F1.
|
|
|
a) |
|
|
|
b) |
|
Figure 3. Inhibitory effects of F1 and piceid on the expression of EBV lytic proteins, including Rta, Zta and EA-D |
Quantification of Cells Expressing EBV Lytic Protein by Fow Cytometry
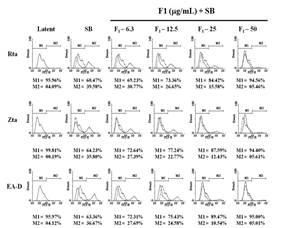
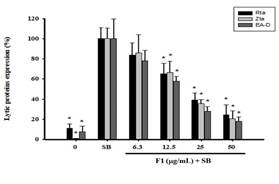
Figure 3 showed that F1 and piceid had the effect of inhibiting the lytic proteins Rta, Zta, and EA-D. Therefore, we further used flow cytometry to quantify cells expressing EBV lytic proteins. P3HR1 cells (6 × 105/mL) were treated with SB for 24 hours, and the percentages of cells expressing Rta, Zta, and EA-D lytic proteins were analyzed by flow cytometry showing the results being 39.6%, 35.8%, and 36.7%, respectively. When treated with an F1 concentration of 25 μg/mL before SB induction, the percentage of cells expressing Rta, Zta, and EA-D decreased to 15.6%, 12.4%, and 10.5%, respectively; when the concentration was increased to 50 μg/mL, it further decreased to 5.5%, 5.6%, and 5.0%, respectively (Figure 4a). The number of cells (M2) expressing Rta, Zta, and EA-D induced by SB was regarded as total expression, and then the data from three independent flow cytometry analyses were quantified. The results showed significant differences between cells treated with SB and 12.5 μg/ml of F1 before SB induction in Rta, Zta, and EA-D protein expression, with only 65.2%, 66.6 %, and 57.8% left, respectively; when the concentration was increased to 25 μg/mL, it decreased to 39.2%, 35.5%, and 28.0%. At 50 μg/mL, the expressions (%) were only 24.3%, 20.7%, and 17.9%, showing that F1 had dose-dependent effects on the inhibition of lytic protein expression. Further assessment on the concentration required for F1 to inhibit 50% of the expressions of Rta, Zta, and EA-D was conducted, and EC50 was 25.9 μg/mL, 25.0 μg/mL, and 19.0 μg/mL, respectively (Figure 4c).
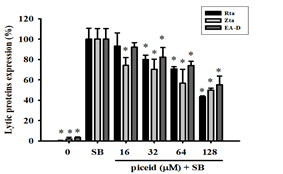
The evaluation of the inhibitory effects of piceid showed that the number of cells expressing Rta, Zta, and EA-D lytic proteins in the SB group was 34.27%, 44.1%, and 34.9%, respectively. At concentrations of 32 μM, 64 μM, and 128 μM, piceid demonstrated the ability to inhibit EBV lytic protein expression. At 128 μM, the percentages of Rta, Zta, and EA-D-positive cells decreased to 14.83%, 22.03%, and 19.27%, respectively (Figure 4b). Cells expressing Rta, Zta, and EA-D lytic proteins were quantified, and the results showed that the number of Zta-positive cells was significantly different from that of the SB control group at a concentration of 16 μM, with a decrease of about 25.8%. There was also a significant reduction in the number of cells expressing Rta and EA-D at a piceid concentration of 32 μM, compared with the SB control group, with a reduction by approximately 20.0% and 17.7%, respectively; at 128 μM, Rta, Zta, and EA-D lytic protein cells were reduced by 56.7%, 50.0%, and 44.8%, respectively. Piceid has a dose-dependent activity on inhibiting EBV lytic protein expression. EC50 values for Rta, Zta, and EA-D expressions were 111.0 μM, 109.1 μM, and 138 μM, respectively (Figure 4d).
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
Figure 4. Quantification of cells expressing EBV lytic proteins, including Rta, Zta, and EA-D treated with F1 and piceid. |
Inhibition of the Transcription of EBV Immediate-Early Genes
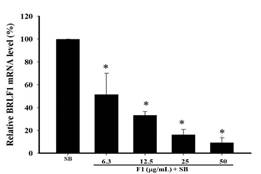
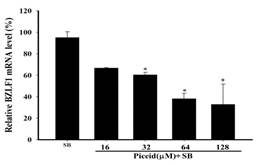
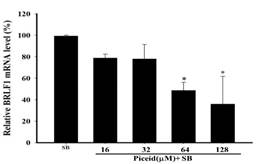
The expression of EBV immediate-early genes, BRLF1, and BZLF1 mRNA, in P3HR1 cells, was analyzed by reverse transcription and quantitative PCR (qPCR). Figure 5a and Figure 5b show that the ethyl acetate subfraction F1 from Polygonum cuspidatum root significantly reduced the transcription of BZLF1 and BRLF1 compared with the SB induction group. The mRNA expression levels of BZLF1 and BRLF1 treated with SB were 97.7 ± 2.1% and 99.8 ± 0.2%, respectively. When 6.3 μg/mL of F1 was added before lytic induction, BZLF1 and BRLF1 expressions were significantly reduced, with levels being only 59.1 ± 4.2% and 51.3 ± 18.7%. The levels of BZLF1 and BRLF1 mRNA were lowered to 21.3 ± 5.65% and 16.5 ± 4.9%, respectively, at 25 μg/mL. When increased to 50 μg/mL, the levels significantly reduced the transcription of BZLF1 and BRLF1 genes, being only 8.6 ± 0.2% and 9.1 ± 4.6%, respectively. Figure 5c and Figure 5d indicate that piceid inhibited EBV lytic gene transcription induced by SB and showed dose effects. When 32 μM and 64 μM piceid were added to the concentration, the expression of BZLF1 mRNA decreased to 60.3 ± 2.3% and 38.1 ± 5.1%, respectively. When the concentration was increased to higher than 64 μM, piceid significantly inhibited the expressions of BRLF1 mRNA; at 64 μM, the expression of BRLF1 mRNA was only 48.5 ± 7.7%.
|
|
|
a) |
|
|
|
b) |
|
|
|
c) |
|
|
|
d) |
|
Figure 5. Effects of F1 and piceid on the EBV BZLF1 and BRLF1 mRNA expression. |
Inhibition of EBV DNA Replication
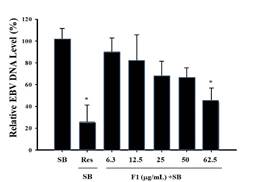
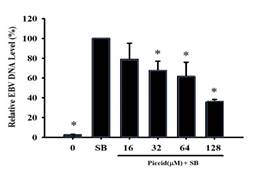
Results from immunoblot and flow cytometry analysis found that F1 and piceid effectively inhibited the expressions of EBV lytic proteins Rta, Zta, and EA-D induced by SB at a non-cytotoxic concentration. We further explored whether F1 and piceid reduce EBV replication. P3HR1 cells were treated with 6.3-62.5 μg/mL of F1 or 16-128 μM piceid before lytic induction. After culturing for two days, EBV DNA in P3HR1 cells was isolated by qPCR to determine the amount (AlHammami et al., 2022; Das et al., 2022; Egwunatum et al., 2022; Gopal & Gurusiddappa, 2022; Roy et al., 2022). From Figure 6a, compared with the group induced by SB alone, F1 at 12.5 μg/mL inhibited EBV DNA replication in the group pretreated with SB followed by the F1 treatment, with a decrease of about 18%. When the concentration was increased to 62.5 μg/mL, there was a significant difference (p<0.05) among groups, with a reduction in EBV DNA replication by 56% and the EC50 being 55.5 μg/mL. In addition, the group pretreated with 55 μM of resveratrol and SB addition, which served as the positive control group, showed a reduction in EBV DNA replication by 74%. Compared with the SB group, at a concentration of 32 μM, piceid was shown to significantly inhibit EBV DNA replication with a reduction of 33%; at the concentrations of 64 μM and 128 μM, the inhibition rates were 38% and 64%, respectively, shown in Figure 6b. The EC50 was 89.9 μM.
|
|
|
a) |
|
|
|
b) |
|
Figure 6. Inhibition Effects of F1 and piceid on EBV DNA replication |
Drugs currently used clinically to treat EBV lytic infection are mostly anti-herpes virus agents, such as acyclovir (ACV), a guanosine analogue, and ganciclovir (GCV). The main mechanism of action is to monophosphorylate ACV and GCV by EBV-encoded protein kinase, and then use the host thymidine kinase to make triphosphorylated ACV and GCV that further inhibits EBV DNA polymerase and replication of viral DNA (Pagano et al., 2018). Although these drugs can inhibit EBV DNA replication, they are highly toxic to patients and show negative effects. They are likely to cause decreased white blood cell or platelet counts in patients and increased side effects such as anemia, skin rash, and abnormal liver function. Therefore, in recent years, there have been numerous studies on natural Chinese herbal medicines aiming to find agents that can effectively inhibit EBV entry into the lytic cycle by environmental factors resulting in subsequent infection or diseases.
Previous literature has confirmed that natural products and Chinese herbal medicines have anti-EBV lytic cycle activity such as EGCG (Chang et al., 2003) and andrographolide (Lin et al., 2008) at a concentration of 100 μM and 14 μM, respectively in P3HR-1 cells that are not cytotoxic and can completely inhibit transcription and expression of EBV lytic proteins in very early stage. Glycyrrhizic acid, at 40 μM, can inhibit early EBV lytic replication with an EC50 of 30 μM (Lin, 2003). Moronic acid inhibits the production of Rta, Zta, and EA-D proteins at 10-20 μM, reduces the expression of BZLF1 and BRLF1 mRNA at 5-15 μM and inhibits the EC50 of Rta at 3.2 μM (Chang et al., 2010).
This study proved that the main component of F1 contained 20.5% of piceid and 10.4% of resveratrol, which can inhibit the expression of EBV lytic proteins, with EC50 of F1 being 23.3 μg/mL and EC50 of piceid being 119.6 μM (46.7 μg/mL). Yiu et al. reported that resveratrol could inhibit EBV lytic proteins with an EC50 of 24 μM (5.5 μg/mL) (Yiu et al., 2010). It is speculated that resveratrol, piceid, and other compounds may be involved in the F1 inhibitory effect on the expression of EBV lytic proteins. Furthermore, it was found that piceid significantly inhibited EBV lytic protein expression, with an EC50 of 119.6 μM. In the cytotoxicity test of P3HR1, CC50 of piceid was 507.9 μM, and the therapeutic index (TI) was 4.2. Piceid and resveratrol have similar structures, with an additional sugar group. In terms of efficacy, resveratrol inhibited 50% of EBV lytic protein expression with an EC50 being 24 μM, CC50 of P3HR1 cells being 137.6 μM, and TI being 5.7. Both substances can inhibit EBV lytic cycle at a nontoxic concentration. In inhibiting the performance of Epstein-Barr virus lytic protein, resveratrol is more effective than piceid. Previous literature pointed out that rutin’s glycosidic bonds can be hydrolyzed through the enzyme system of bacteria in the gastrointestinal tract, and then transform into quercetin and then enter the cell to exhibit better biological activity (Yang et al., 2012). It is further speculated that piceid and resveratrol may also have the same effects as rutin and quercetin; piceid may exhibit a greater inhibitory effect on the EB virus lytic cycle after biological metabolism than resveratrol.
The possible mechanism for the resveratrol and piceid to activate the deacetylation activity of sirtuin is by inhibiting p38 MAPK, ERK, and JNK transduction that affects AP-1 and ATF2 activation, preventing activation of EBV immediate-early genes, and thereby inhibiting EBV lytic cycle (Yiu et al., 2010).
Conclusion
Our study results demonstrate that F1 and piceid could inhibit lytic gene transcription, prevent EBV lytic cycle, and reduce viral DNA replication. The ability of F1 and piceid to affect EBV reactivation has the potential to be used to develop anti-EBV drugs.
Acknowledgments: None
Conflict of interest: None
Financial support: This work was supported by research grants NSC100-2320-B-041-004-MY3 and MOST 103-2320-B-041 -002 from the Ministry of Science and Technology, Taiwan, and CLFHR10105 and CLFHR10302 from Chi Mei Medical Center, Liouying, Taiwan.
Ethics statement: None
Aiche, S., Smail, F., Chikhaoui, M., & Abdelhadi, S. A. (2022). Influence of age, parity, and fetuses’ number on blood parameters in rembi ewes during late pregnancy in semi-arid zone, Algeria. World Journal of Environmental Biosciences, 11(2), 26-33. doi:10.51847/jD3o3GsDHr
AlHammami, Q. S., AlThobaiti, M. S., AlDhasi, A. A., AlMutairi, Y. A. A., Kabrah, L. K., AlShehri, A. M., Alshwamen, M. S. M., & AlSulimi, L. (2022). Updates on the role of interventional radiology in ischemic patients undergoing aortic valve replacement: systematic review. World Journal of Environmental Biosciences, 11(3), 30-35. doi:10.51847/Mpx8ibFID4
Almaghrabi, S. Y., & Hussein, K. S. (2022). Microbial profile of women with preterm premature rupture of membrane in Saudi Arabia: a retrospective study. World Journal of Environmental Biosciences, 11(2), 43-47. doi:10.51847/0ij9xVzYK7
Anderson, A. G., Gaffy, C. B. Weseli, J. R., & Gorres, K. L. (2019). Inhibition of Epstein-Barr Virus Lytic Reactivation by the Atypical Antipsychotic Drug Clozapine. Viruses, 11(5), 450. doi:10.3390/v11050450
Andrei, G., Trompet, E., & Snoeck, R. (2019). Novel Therapeutics for Epstein⁻Barr Virus. Molecules, 24(5), 997. doi:10.3390/molecules24050997
Benedetti, F., Sorrenti, V., Buriani, A., Fortinguerra, S., Scapagnini, G., & Zella, D. (2020). Resveratrol, Rapamycin and Metformin as Modulators of Antiviral Pathways. Viruses, 12(12), 1458. doi:10.3390/v12121458
Chandra, S., & Meel, R. K. (2022). A systematic comparative study of morinda tinctoria and vitex negundo for their anti-ulcerogenic potential. World Journal of Environmental Biosciences, 11(1), 45-52. doi:10.51847/aNF9QSYDRo
Chang, F. R., Hsieh, Y. C., Chang, Y. F., Lee, K. H., Wu, Y. C., & Chang, L. K. (2010). Inhibition of the Epstein-Barr virus lytic cycle by moronic acid. Antiviral Research, 85(3), 490-495. doi:10.1016/j.antiviral.2009.12.002
Chang, L. K., Wei, T. T., Chiu, Y. F., Tung, C. P., Chuang, J. Y., Hung, S. K., Li, C., & Liu, S. T. (2003). Inhibition of EBV lytic cycle by (-)-epigallocatechin gallate. Biochemical and Biophysical Research Communications, 301(4), 1062-1068. doi:10.1016/S0006-291X(03)00067-6
Chen, C. C., Chang, K, C., Medeiros, L. J., & Lee, J. Y. (2020). Hydroa Vacciniforme and Hydroa Vacciniforme-Like Lymphoproliferative Disorder: A Spectrum of Disease Phenotypes Associated with Ultraviolet Irradiation and Chronic Epstein-Barr Virus Infection. International Journal of Molecular Sciences, 21(23), 9314. doi:10.3390/ijms21239314
Das, S. K., Karan, S., & Sen, K. (2022). Biodiversity of avifauna in chilkigarh, jhargram, West Bengal, India. World Journal of Environmental Biosciences, 11(3), 8-13. doi:10.51847/jNtkP7dkxS
Dochi, H., Kondo, S., Murata, T., Fukuyo, M., Nanbo, A., Wakae, K., Jiang, W. P., Hamabe-Horiike, T., Tanaka, M., Nishiuchi, T., et al. (2022). Estrogen induces the expression of EBV lytic protein ZEBRA, a marker of poor prognosis in nasopharyngeal carcinoma. Cancer Science, 113(8), 2862-2877. doi:10.1111/cas.15440
Dugan, J. P., Haverkos, B. M., Villagomez, L., Martin, L. K., Lustberg, M., Patton, J., Martin, M., Huang, Y., Nuovo, G., Yan, F., et al. (2018). Complete and Durable Responses in Primary Central Nervous System Posttransplant Lymphoproliferative Disorder with Zidovudine, Ganciclovir, Rituximab, and Dexamethasone. Clinical Cancer Research, 24(14), 3273-3281. doi:10.1158/1078-0432.CCR-17-2685
Egwunatum, A. E., Uyovbisere, E., & Umeh, L. C. (2022). Effect of forest-incubated composts on crude-oil soils for Zea mays, L. cultivation in Delta State, Nigeria. World Journal of Environmental Biosciences, 11(3), 14-20. doi:10.51847/j5Pyls0seh
Fayiah, M., Tulcan, R. X. S., Bockarie, R., Saccoh, S., Singh, S., & Fayiah, M. S. (2022). Sierra Leone rainfall and drought dynamics from 1990-2020: a holistic perspective. World Journal of Environmental Biosciences, 11(2), 34-42. doi:10.51847/8Wz28ID8Mn
Gopal, V., & Gurusiddappa, L. H. (2022). Influence of jeevamrutha (fermented liquid manure) on growth and yield parameters of tomato (Solanum LycopersicumL.). World Journal of Environmental Biosciences, 11(3), 1-7. doi:10.51847/WFD516GS8o
Heslop, H. E. (2020). Sensitizing Burkitt lymphoma to EBV-CTLs. Blood, 135(21), 1822-1823. doi:10.1182/blood.2020005492
Jangra, S., Yuen, K. S., Botelho, M. G., & Jin, D. Y. (2019). Microorganisms, 7(6), 183. doi:10.3390/microorganisms7060183
Karami, A., Fakhri, S., Kooshki, L., & Khan, H. (2022). Polydatin: Pharmacological Mechanisms, Therapeutic Targets, Biological Activities, and Health Benefits. Molecules, 27(19), 6474. doi:10.3390/molecules27196474
Koike, R., Nodomi, K., Watanabe, N., Ogata, Y., Takeichi, O., Takei, M., Kaneko, T., Tonogi, M., Kotani, A. I., & Imai, K. (2020). Butyric Acid in Saliva of Chronic Periodontitis Patients Induces Transcription of the EBV Lytic Switch Activator BZLF1: A Pilot Study. In Vivo, 34(2), 587-594. doi:10.21873/invivo.11811
Li, H., Li, Y., Hu, J., Liu, S., Luo, X., Tang, M., Bode, A. M., Dong, Z., Liu, X., Liao, W., et al. (2021). (−)-Epigallocatechin-3-Gallate Inhibits EBV Lytic Replication via Targeting LMP1-Mediated MAPK Signal Axes. Oncology Research, 28(7), 763-778. doi:10.3727/096504021X16135618512563
Lin, J. C. (2003). Mechanism of action of glycyrrhizic acid in inhibition of Epstein-Barr virus replication in vitro. Antiviral Research, 59(1), 41-47. doi:10.1016/S0166-3542(03)00030-5
Lin, T. P., Chen, S. Y., Duh, P. D., Chang, L. K., & Liu, Y. N. (2008). Inhibition of the EBV lytic cycle by andrographolide. Biological Pharmaceutical Bulletin, 31(11), 2018-2023. doi:10.1248/bpb.31.2018
Murata, T., Sugimoto, A., Inagaki, T., Yanagi, Y., Watanabe, T., Sato, Y., & Kimura, H. (2021). Molecular Basis of Epstein-Barr Virus Latency Establishment and Lytic Reactivation. Viruses, 13(12), 2344. doi:10.3390/v13122344
Pagano, J. S., Whitehurst, C. B., Andrei, G. (2018). Antiviral Drugs for EBV. Cancers (Basel), 10(6), 197. doi:10.3390/cancers10060197
Pimple, N. S. (2022). Virtual population analysis and recruitment pattern of Osteobrama vigorsii (Sykes, 1839) from Nira River, Bhor Maharashtra. World Journal of Environmental Biosciences, 11(1), 53-59. doi:10.51847/QslcwlezoR
Roy, S., Laha, I., Ray, D., & Choudhury, L. (2022). Influence of climate change & environmental toxicants on epigenetic modifications. World Journal of Environmental Biosciences, 11(3), 21-29. doi:10.51847/jku3EDOAkt
Shao, Q., Liu, T., Wang, W., Liu, T., Jin, X., & Chen, Z. (2022). Promising Role of Emodin as Therapeutics to Against Viral Infections. Frontiers in Pharmacology, 13, 902626. doi:10.3389/fphar.2022.902626
Wong, Y., Meehan, M. T., Burrows, S. R., Doolan, D. L., & Miles, J. J. (2022). Estimating the global burden of Epstein-Barr virus-related cancers. Journal of Cancer Research and Clinical Oncology. 148, 31-46. doi:10.1007/s00432-021-03824-y
Wu, C. C., Chen, M. S., Cheng, Y. J., Ko, Y. C., Lin, S. F., Chiu, I. M., & Chen, J. Y. (2019). Emodin inhibits EBV reactivation and represses NPC tumorigenesis. Cancers, 11(11), 1795-1812. doi:10.3390/cancers11111795
Wu, X., Li, Q., Feng, Y., & Ji, Q. (2018). Antitumor Research of the Active Ingredients from Traditional Chinese Medical Plant Polygonum Cuspidatum. Evidence-Based Complementary and Alternative Medicine, 2018, 2313021. doi:10.1155/2018/2313021
Yang, J., Qian, D., Jiang, S., Shang, E., Guo, J., & Duan, J. (2012). Identification of rutin deglycosylated metabolites produced by human intestinal bacteria using UPLC-Q-TOF/MS. Journal of Chromatography B, 898(1), 95-100. doi:10.1016/j.jchromb.2012.04.024
Yiu, C. Y., Chen, S. Y., Chang, L. K., Chiu, Y. F., & Lin, T. P. (2010). Inhibitory effects of resveratrol on the EBV lytic cycle. Molecules, 15(10), 7115-7124. doi:10.3390/molecules15107115
Yiu, C. Y., Chen, S. Y., Yang, T. H., Chang, C. J., Yeh, D. B., Chen, Y. J., & Lin, T. P. (2014). Inhibition of Epstein-Barr virus lytic cycle by an ethyl acetate subfraction separated from Polygonum cuspidatum root and its major component, emodin. Molecules, 19(1), 1258-1272. doi:10.3390/molecules19011258
Yiu, C. Y., Chiu, Y. J., & Lin, T. P. (2021). The Ethyl Acetate Subfraction of Polygonum cuspidatum Root Containing Emodin Affect EBV Gene Expression and Induce EBV-Positive Cells Apoptosis. Biological Pharmaceutical Bulletin, 44(12), 1837-1842. doi:10.1248/bpb.b21-00508
Yun-Ting, Z., Xiao, H., Yun-Zhong, C., Jun-de, L. I., & Kun, Y. U. (2020). Chemical constituents and their biosynthesis mechanisms of Polygonum cuspidatum. Zhongguo Zhong Yao Za Zhi, 45(18), 4364-4372. doi:10.19540/j.cnki.cjcmm.20200525.201
Zhang, S., Zhu, Y., Jin, Y., Sun, H., Wang, W., & Zhan, L. (2021). Difference between Acyclovir and Ganciclovir in the Treatment of Children with Epstein–Barr Virus-Associated Infectious Mononucleosis. Evidence-Based Complementary and Alternative Medicine, 2021, 8996934. doi:10.1155/2021/8996934