A Review on Biotechnological Approaches for the Production of Polyhydroxyalkanoates
|
Aluru Ranganadhareddy* Department of Biotechnology, VFSTR Deemed to be University, Guntur, India 522213.
|
*E-mail: [email protected]
Abstract
Producers of Polyhydroxyalkanoates (PHA) are reported to stay at different ecological niches that are exposed naturally or accidentally to high organic matter or growth-limiting conditions like wastes from pulp and paper mills, wastes from dairy, wastes from agriculture, sites contaminated by hydrocarbons, treatment plants releasing activated sludges, rhizosphere, and effluents from industries. Extracellular secondary products such as rhamnolipids, polymeric substances as well as biohydrogen gas are also produced by a few among them. These microbes can produce valuable bioproducts like PHA with the simultaneous usage of waste materials having various origins as substrate and for this reason, these types of microbes are industrially important candidates in regard to production. An economical method could be the separation of the secondary products (intracellular and extracellular) by the implementation of an integrated system. Different microorganisms living under various environmental conditions that stimulate the microorganisms to store carbon as polyhydroxyalkanoates granules have been discussed in this review. We have also discussed various metabolites that are produced concurrently with PHA.
Keywords: Polyhydroxyalkanoates, Microbes, Biohydrogen, Rhamnolipids
Introduction
Suitable places for microorganisms to perform individual roles and communicate with each other to form an ecosystem are called ecological niches. Due to natural and man-made activities, biotic and abiotic altercations influence the ecosystem. To put together a thriving existence, microbes obtain their sustenance from various behavioral changes in the environment. Among such responses of microbes living in various ecological niches towards experienced stress, PHA accumulation is one Figure 1.
|
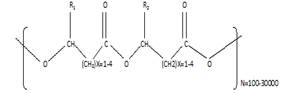
|
|
Figure 1. Polyhydroxyalkanoates structure (R1 and R2 are alkyl groups C-C) (Ranganadhareddy, 2022) |
Ecological niches such as delta deposits, aquatic microbial mats, root microbiome, groundwater deposits, and artificial ecosystems which have shifting nutrient contents support the organisms highly engaged in the accumulation of PHA to meet the energy requirements for metabolism during the period of starvation. This concept can be executed industrially with an acceptable production process to reduce the commercial cost of biopolymers (Reddy et al., 2019). Predominant ecological conditions and phenotype traits of PHA producers are to be understood clearly. Harmful, growth hindering, and unknown environmental conditions are created by waste disposal (Ranganadhareddy, 2022). Even during such physiological strain, PHA producers are reported to reside with properties such as aromatic compounds, degrading dyes, as well as residual organic matter in industrial discharge (Ravi Teja et al., 2020). This improbable behavioral character may be regarded towards the aspect of industrial production as well as various issues connected with the withdrawal of pollutants such as pesticides, dyestuffs, PHA, etc. Various wastes such as wastes from dairy (Indira et al., 2016), effluents from textile (Ranganadhareddy 2022), distillery effluent (Zhang et al., 2021), wastes from oil, paper mills (Reddy et al., 2018), residues of animals (Maier et al., 2017), starchy wastes, wastes from agriculture (Ferreira et al., 2018) and by-products of biodiesel wastes (Andler et al., 2021) are being tried by researchers for use in the production of PHA, with apprehension about the growing load of wastes from industries. With an excess of carbon under nutrient-limiting conditions, the production of PHA occurs (Reddy et al., 2019). This produced PHA is then stored as inclusion bodies which is a polymerized form (Reddy et al., 2017). Various aspects of the production of bioplastic with the use of activated sludge as the producer of PHA (Lamberti et al., 2020) enriched by aerobic dynamic feeding as well as the alternating aerobic and anaerobic system in sequencing batch reactor and fermented effluents showing high values of Biological Oxygen Demand and Chemical Oxygen Demand as substrates have been contributed over the last few years (Ohnishi et al., 2022). Without a doubt, it is understood that the requirement of an established designing of operational framework to answer the issue related to aseptic conditions, artificial medium composition, and continuance of pure cultures, as well as the high cost of the product, is ruled out by the production of PHA from a mixed culture, enhanced phosphate accumulating organisms, enhanced glycogen accumulating organisms and low or high salt conditions inhabited by Archaea (Mitra et al., 2020). During various environmental stresses such as nutrient-limiting conditions, cold treatment, ethanol, hydrogen peroxide, UV-irradiation, or osmotic shocks, it has been reported that stored co-polyesters PHBHHx in Aeromonas hydrophila 4AK4 can survive (Obruca et al., 2022). The practicality to pull through during these harsh conditions which are not favorable for the usual growth of cells is enhanced by the production of PHA. It has been proved that certain wastes such as seafood cannery that is rich in ammonia, phosphates, and lipids are virulent to the microbes. Here, the accession of PHB has an advantage, as it works as both a stored product as well as an alternative to burning surplus fatty acids to cope with the existing stress (Kumar et al., 2020). A possible advantage is using the intracellularly stored carbon source at a later phase to support the activities of the cell. This is of notable importance as it is a commercially valuable secondary product. A few microorganisms also synthesize extracellular metabolites such as alginate oligosaccharides, biohydrogen, rhamnolipids, and extracellular polymeric substances, as well as intracellular production of PHA as inclusion bodies (Ranganadha et al., 2021). It has also been reported by studies, how PHA production and PHA polymer composition are modulated by the composition, concentration, and uptake of the substrate. The production and composition of PHA have also reportedly been affected by pH and temperature (Reddy et al., 2018). Therefore, comprehension of the PHA synthesis, as well as the related kinetics in regard to the concurrent synthesis of other secondary products, can provide a platform for a cost-effective and efficient manufacturing unit.
Common Pathways for the Synthesis of Polyhydroxyalkanoates
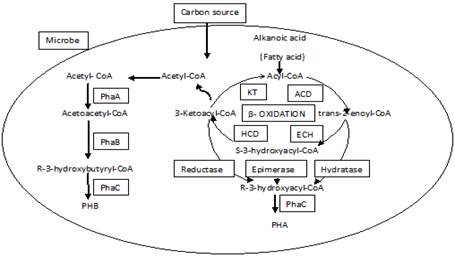
Through the Calvin Benson cycle, atmospheric CO2 is converted into PHA, which is converted to Acetyl-CoA through glycolysis. Acetyl-CoA is converted into the most prevalent PHA, PHB by three enzymatic reactions (Syahirah et al., 2021). Two Acetyl-CoA molecules are combined into Acetoacetyl-CoA by the first enzyme, PhaA (PHA-specific ß-ketothiolase). PhaB (Acetoacetyl-CoA reductase) is the second enzyme that helps in the formation of hydroxybutyryl-CoA by the reduction of Acetoacetyl-CoA. Finally, a growing PHB molecule is added with hydroxybutyryl-CoA fatty acid monomer via an ester bond by PhaEC (PHB synthase) (Mohapatra et al., 2017). Figure 2 shows the reaction scheme. There are no significant differences between heterotrophic and phototrophic organisms when it comes to the synthesis of PHB (Zhang et al., 2020). However, in heterotrophs (Cupriavidus nectator), PhaA, PhaB, and PhaEC genes are located in a single operon while in cyanobacteria (SynechocystisPCC6803), two separate operons are required which are approximately 500Kbp apart. First loci are thought to contain PhaA and PhaB genes, each of which is putatively co-expressed. PhaEC is a heterodimer made up of PhaE and PhaC and is present at the second locus.
|
|
|
Figure 2. Biosynthetic pathway of Polyhydroxybutyrate in microbes (Ranganadha et al., 2021) |
PHA-Producing Bacteria Along with their Ecological Niches
Hydrocarbon Degrading Bacteria as PHA Producers
The existence of xenobiotic components acts as environmental stress to divert the inhabiting organisms’ physiological reactions to synthesize PHA (Monroy & Buitrón, 2020). A true requirement for the production of PHA by cells is created by areas contaminated by oil that have higher carbon concentrations (84%) with a less content of nitrogen (>1%) (Chen et al., 2019). Various bacterial strains inherent in the genera Sphingobacterium, Yokenella, Acinetobacter, Pseudomonas, Caulobacter, Burkholderia, Ralstonia, and Brochothrix have been isolated from sites contaminated with oil that can produce PHA while degrading oil. In an experiment, it was reported that Cupriavidus necator DSM 545can produce 62% PHB of cell dry weight when it is given growth impeding material like waste glycerol a byproduct from the biodiesel industry as a source of carbon. Thus, for the production of valuable bioproducts, it is of colossal need to avert inebriation and survey the potential of microbes. In one case, it has been reported that a deviant aquatic bacterium AlcanivoraxborkumensisSK2 when allowed to grow on aliphatic hydrocarbon, deposits PHB extracellularly. A reduction in the cost of the manufacturing unit along with efficient removal of unwanted substances can be achieved by the establishment of a bio-plant where unwanted toluene as the sole source of carbon ensures that Rhodococcus aetherivorans IARI produces copolymer P(3HB-co-HV) along with triacylglycerols (Filiz et al., 2021; Rosenboom et al., 2022). This proved to be a probable candidate for the altering of petrochemical waste into valuable substances such as PHA (AlZahrani, 2023).
Halophilic Bacteria as PHA Producers
Since it has been reported that archaea live in hot springs, oceans, marshlands, salt lakes, etc., they are considered extremophiles. Archaea also produce PHA, reportedly. Salts sustain the growth of these organisms. 5% is required for optimal growth and they tolerate salt NaCl (w/v) of at least 10%. (Koller, 2019) Accumulation of PHA by archaea had been reported for the first time from the Dead Sea and named Halobacterium marismortui in the year 1970. It had been analyzed by the technique of freeze-fracture. PHB is produced in limiting conditions of nutrients and excess sources of carbon by extremely halophilic archaebacteria. Halomonas venusta, a moderate Halophile produces a higher amount of PHB of up to 69% of CDW when it is allowed to grow on glucose as substrate (Gao & Zhang, 2014; Kassari et al., 2020). Pretreatment converts organic substances from a more complex form into simpler forms that are easily consumable. Maltose from starch hydrolysates is preferably utilized by H. boliviensisLC1. It has been found that PHA production is enhanced by oxygen limitation. Most importantly, it has been reported that vinasses (secondary product of ethanol industries) used as substrates for extremely halophilic archaea Haloarculamaris mortui leads to the production of PHA under shake flask conditions (Tan et al., 2021).
Plant Growth Promoting Rhizobia bacteria as PHA Producers
Extracellular metabolites which enhance the root and plant growth are secreted from microbes harbored in the soil surrounding the plant roots. This is called the rhizosphere. Recycling of nutrients takes place in the rhizosphere which serves as a hotspot for microbial interaction due to the release of nutrients into the surrounding rhizosphere by root exudates. Along with the effect of antagonists and PGPR, it has been interpreted that the soil surrounding the roots also has reservoirs for accumulators of PHA that are hidden or untapped. A rhizobacterium that promotes the growth of the plant and is being progressively utilized in agriculture at a commercial scale is Azospirillum brasilense. Recent research has elucidated various key properties of A. brasilense which provide the ability to adapt to the habitat of the rhizosphere as well as to promote the growth of the plant. The ability to sustain variation in phenotype as well as the production of the auxin indole-3-acetic acid, carotenoids, a range of cell surface components, and nitric oxide are some of these key properties. It was reported that the variants of Azospirillum brasilense genes phaC and phaZ were less resilient to unfavorable conditions like heat, osmotic pressure, UV-radiation, presence of H2O2, and desiccation when compared to wild type (Ranganadha & Chandrasekhar, 2021). PHA production has characteristics that are advantageous to the promotion of plant growth, enrichment of colonization of roots, survivability, motility, chemotaxis as well as multiplication of cells. A. brasilense inoculants’ reliability, efficiency, and shelf life can be improved by the production of PHA. When compared to non-rhizosphere soil, the rhizosphere reportedly has low production of PHA based on cultivation-dependent methods (Kumar & Kim, 2018; Li & Singh, 2021). Production of PHA was more for wheat, oilseed, rape, and sugar-beet rhizosphere as concluded by cultivation-independent and molecular techniques. More PHA producers are harbored in the roots of the plants producing oil and carbohydrates as supported by the fact that inorganic nutrients are a limited and abundant source of carbon contained in the root exudates. The accumulation of PHB was seen to be a numerical character that is distinctively controlled by different genes in rhizobia (Kumar & Kim, 2018; Wu et al., 2022).
PHA and Extracellular Polymeric Substances
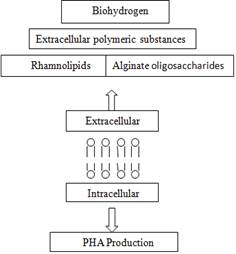
When subjected to unfavorable growth conditions, extracellular polymeric substances protect and supply energy to cells (Alharbi et al., 2023; Alziyadi et al., 2023; Ilias et al., 2023; Suragimath et al., 2023; Voiţă-Mekereş et al., 2023). When the substrate is limited, a mixture of high molecular polymers serves as suppliers of carbon units. When organisms are under starvation conditions, an extracellular product EPS and an intracellular product PHA are produced. In Ralstonia eutropha ATCC17699, Rhizobium meliloti, Azospirillum brasilense, and Anabaena cylindrica, simultaneous production of EPS and PHA were reported Figure 3.
|
|
|
Figure 3. Simultaneous production of intracellular and extracellular products (Baljeet et al., 2014) |
It was also reported about the positive and negative effects of nitrogen concentration on EPS production (Gaignard et al., 2019). In Azotobacter chroococcum in nitrogen limiting conditions, EPS production was initiated with fructose and glucose as efficient carbon sources, whereas in Ralstonia eutropha ATCC17699 an increase in nitrogen and glucose supply enhanced the production of EPS in batch cultures (Kumar & Kim, 2018; Kardile & Shirsat, 2020). For the PHB synthesis, the decomposition of EPS served as a nitrogen source. On the other hand, when the external carbon source gets depleted, PHB decomposition served as a supplier of carbon sources (Abou-assy et al., 2023; Elssayed et al., 2023; Mady et al., 2023; Yousif et al., 2023).
PHA and Biohydrogen
A promising technology attributed to its purity and fewer energy requirements is the biological production of Hydrogengas (Guerra et al., 2019). The production of hydrogen and PHA under nutrient-limited conditions is reported to occur in many purple non-sulfur bacteria like Rhodospirillum rubrum, Rhodobacter sphaeroides, Rhodopseudomonas palustris, and Bacillus sp. PHB synthesis is conducive to Hydrogen production when strain WP3-5 is in stressful conditions although PHB synthesis does compete with hydrogen production according to the literature (Ranaivoarisoa et al., 2019). To reduce the cost of bioproducts, an efficient light-dependent process is the production of Hydrogen gas which occurs in the first stage (nitrogen-limited) and subsequent transfer of culture to the second stage (phosphorus-limited) where PHB production occurs.When grown on glucose in two-stage systems, Hydrogen gas in the range of 1.67 to1.92mol H2 mol−1 glucose−1 and 11.5% of CDW of PHB was produced by some non-photosynthetic bacterium like Bacillus thuringiensis UGU45 and Bacillus cereusEGU44 (Patel et al., 2011). According to a study, when subjected to extra environmental stress conditions like suboptimal pH value, the Hydrogen producing ability of Rhodopseudomonas palustris WP3-5 bacterium is retained by stored PHB (Ranaivoarisoa et al., 2019).
Conclusion
PHA, a biodegradable material stored by microorganisms, is an alternative to petroleum-derived plastics. The environment can be examined to investigate different PHB accumulating strains. Polyhydroxtbutyratye contributes to the creation of a sustainable environment by replacing non-biodegradable synthetic polymers. To compete with the thermoplastics-dominated industry, it is critical to develop low-cost biodegradable polymers. Biopolymers' important characteristics, such as biodegradable and biocompatible nature, have made them suitable for use in the food packaging industry, aqua industry biomedical industry, and antifouling. As a result of this report, it is possible to conclude that PHB is a durable class of biopolymers that may be utilized to improve the design of new applications in a variety of industries.
Acknowledgments: None
Conflict of interest: None
Financial support: None
Ethics statement: None
Abou-assy, R. S., Aly, M. M., Amasha, R. H., Jastaniah, S., Alammari, F., & Shamrani, M. (2023). Carbapenem resistance mechanisms, carbapenemase genes dissemination, and laboratory detection methods: a review. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 123-138. doi:10.51847/wqUTf4VfuO
Alharbi, H. M., Alqahtani, T., Batubara, A., Alshaer, A., Alqurashi, B., Bahwairth, L., Khawaji, H., & Almohammadi, A. M. (2023). Enhancing the dissolution of oral dasatinib tablets using zein–hydroxypropyl methylcellulose solid dispersions. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 139-148. doi:10.51847/crBI2n6KLG
AlZahrani, S. G. (2023). Healthy schools framework in Saudi Arabia: a narrative review. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 110-115. doi:10.51847/lJ2Fbaa0Vm
Alziyadi, S. H., AlNemari, S. M., AlOsaimi, M. M., AlSufyani, M. E., AlReefi, T. M., AlShanbari, S. F., & Koursan, S. M. S. (2023). Oral contraceptive use and its association with depression among women in Taif, Saudi Arabia. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 32-40. doi:10.51847/6PmM8jcQwn
Andler, R., Valdés, C., Urtuvia, V., Andreeßen, C., & Díaz-Barrera, A. (2021). Fruit residues as a sustainable feedstock for the production of bacterial polyhydroxyalkanoates, Journal of Cleaner Production, 307, 127236. doi:10.1016/j.jclepro.2021.127236
Baljeet, S. S., Anita, G., & Pardeep, K. (2014). Biotechnological Production of Polyhydroxyalkanoates: A Review on Trends and Latest Developments. Chinese Journal of Biology, 18. doi:10.1155/2014/802984
Chen, F., Li, X., Zhu, Q., Ma, J., Hou, H., & Zhang, S. (2019). Bioremediation of petroleum-contaminated soil enhanced by aged refuse. Chemosphere, 222, 98-105. doi:10.1016/j.chemosphere.2019.01.122
Elssayed, A., AlRgaiba, R. I., AlZalbani, M. K., Hassan, M. R. J., AlMalki, K. H., AlGhannam, A. A., AlMudayfir, Z. F., Mohamed, H. A. A., Sheikh, M. M., AlGhamdi, A. A., et al. (2023). Review on Diagnosis and Management Approach of Multiple Sclerosis. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 100-105. doi:10.51847/GJcJdSPaJm
Ferreira, F. V., Dufresne, A., Pinheiro, I. F., Souza, D. H. S., Gouveia, R. F., Mei, L. H. I., & Lona, L. M. F. (2018). How do cellulose nanocrystals affect the overall properties of biodegradable polymer nanocomposites: A comprehensive review. European Polymer Journal, 108, 274-285. doi:10.1016/j.eurpolymj.2018.08.045
Filiz, A. K., Joha, Z., & Yulak, F. (2021). Mechanism of anti-cancer effect of β-glucan on SH-SY5Y cell line. Bangladesh Journal of Pharmacology, 16(4), 122-128. doi:10.3329/bjp.v16i4.54872
Gaignard, C., Laroche, C., Pierre, G., Dubessay, P., Delattre, C., Gardarin, C., Gourvil, P., Probert, I., Dubuffet, A., & Michaud, P., (2019). Screening of marine microalgae: Investigation of new exopolysaccharide producers. Algal Research, 44, 101711. doi:10.1016/j.algal.2019.101711.
Gao, S., & Zhang, L. H. (2014). The synthesis of poly-β-hydroxybutyrate by moderately halophilic bacteria Halomonas venusta. In Advanced Materials Research (Vol. 1033, pp. 306-310). Trans Tech Publications Ltd. doi:10.4028/www.scientific.net/AMR.1033-1034.306
Guerra, O. J., Eichman, J., Kurtz, J., & Hodge, B. M. (2019). Cost competitiveness of electrolytic hydrogen. Joule, 3(10), 2425-2443. doi:10.1016/j.joule.2019.07.006
Ilias, T. I., Pantea, V. A., Căiță, G. A., & Pantea-Roșan, L. R. (2023). Investigating the effect of physical activity and exercise on serum leptin levels. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 163-169. doi:10.51847/Y9VAMeiiDU
Indira, M., Venkateswarulu, T. C., Chakravarthy, K., Reddy, A. R., Babu, D. J., & Kodali, V. P. (2016). Morphological and biochemical characterization of exopolysaccharide producing bacteria isolated from dairy effluent. Journal of Pharmaceutical Sciences and Research, 8(2), 88-91.
Kardile, D., & Shirsat, M. (2020). Synthesis and in vitro evaluation of coupled mercaptobenzimidazole derivatives used as a potent biological agent. International Journal of Pharmaceutical and Phytopharmacological Research, 10(1), 127-133.
Kassari, H., Dehghani, R., Kasiri, M., Dehghani, M., & Kasiri, R. (2020). A review on the reappearance of Crimean-Congo hemorrhagic fever, a tick-borne nairovirus. Entomology and Applied Science Letters, 7(1), 81-90.
Koller, M. (2019). Polyhydroxyalkanoate biosynthesis at the edge of water activitiy-haloarchaea as biopolyester factories. Bioengineering, 6(2), 34. doi:10.3390/bioengineering6020034
Kumar, P., & Kim, B. S. (2018). Valorization of polyhydroxyalkanoates production process by co-synthesis of value-added products. Bioresource Technology, 269, 544-556. doi:10.1016/j.biortech.2018.08.120
Kumar, V., Kumar, S., & Singh, D. (2020). Microbial polyhydroxyalkanoates from extreme niches: Bioprospection status, opportunities and challenges. International Journal of Biological Macromolecules, 147, 1255-1267. doi:10.1016/j.ijbiomac.2019.09.253
Lamberti, F. M., Román-Ramírez, L. A., & Wood, J. (2020). Recycling of bioplastics: routes and benefits. Journal of Polymers and the Environment, 28(10), 2551-2571. doi:10.1007/s10924-020-01795-8
Li, F., & Singh, A. I. (2021). Recent advancements to enhance the therapeutic efficacy of antiepileptic drugs. Acta Pharmaceutica, 71(4), 527-544. doi:10.2478/acph-2021-0041.
Mady, M., Arishi, H. A., AlOtaibi, T. N., AlFadhel, A. K., AlRajhi, H. M., Jaafari, F. A., Soliman, S. A., AlShehri, A. H., AlNasser, F. A., AlHomaidhi, S. M., et al. (2023). Literature review on diagnostic and management approach of anodontia. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 106-109. doi:10.51847/MLRKb4kzVC
Maier, S., Szerencsits, M., Narodoslawsky, M., Ismail, I. M. I., & Shahzad, K. (2017). Current potential of more sustainable biomass production using eco-efficient farming practices in Austria. Journal of Cleaner Production, 155(1), 23-27. doi:10.1016/j.jclepro.2016.09.037
Mitra, R., Xu, T., Xiang, H., & Han, J. (2020). Current developments on polyhydroxyalkanoates synthesis by using halophiles as a promising cell factory. Microbial Cell Factories, 19(1), 1-30. doi:10.1186/s12934-020-01342-z
Mohapatra, S., Maity, S., Dash, H. R., Das, S., Pattnaik, S., Rath, C. C., & Samantaray, D. (2017). Bacillus and biopolymer: prospects and challenges. Biochemistry and Biophysics Reports, 12, 206-213. doi:10.1016/j.bbrep.2017.10.001
Monroy, I., & Buitrón, G. (2020). Production of polyhydroxybutyrate by pure and mixed cultures of purple non-sulfur bacteria: A review. Journal of Biotechnology, 317, 39-47. doi:10.1016/j.jbiotec.2020.04.012
Obruča, S., Dvořák, P., Sedláček, P., Koller, M., Sedlář, K., Pernicová, I., & Šafránek, D. (2022). Polyhydroxyalkanoates synthesis by halophiles and thermophiles: Towards sustainable production of microbial bioplastics. Biotechnology Advances, 58, 107906. doi:10.1016/j.biotechadv.2022.107906
Ohnishi, A., Hasegawa, Y., Fujimoto, N., & Suzuki, M. (2022). Biohydrogen production by mixed culture of megasphaera elsdenii with lactic acid bacteria as lactate-driven dark fermentation. Bioresource Technology, 343, 126076. doi:10.1016/j.biortech.2021.126076
Patel, S. K., Singh, M., & Kalia, V. C. (2011). Hydrogen and polyhydroxybutyrate producing abilities of Bacillus spp. from glucose in two stage system. Indian Journal of Microbiology, 51(4), 418-423. doi:10.1007/s12088-011-0236-9
Ranaivoarisoa, T. O., Singh, R., Rengasamy, K., Guzman, M. S., & Bose, A. (2019). Towards sustainable bioplastic production using the photoautotrophic bacterium Rhodopseudomonas palustris TIE-1. Journal of Industrial Microbiology and Biotechnology, 46(9-10), 1401-1417. doi:10.1007/s10295-019-02165-7
Ranganadha, A. R., & Chandrasekhar, Ch. (2021). Production of polyhydroxybutyrate from marine source- A Review. Indian Journal of Ecology, 48(6), 1829-1836.
Ranganadha, A. R., Sravani, K., Sanjana, N., & Chandrasekhar, Ch. (2021). Production of biopolymer from bacteria – A Review. Environmental and Earth Sciences Research Journal, 8(2), 91-96. doi:10.18280/eesrj.080205
Ranganadhareddy, A. (2022). A Review on Production of Polyhydroxyalkanoates in Microorganisms. Journal of Biochemical Technology, 13(1), 1-6. doi:10.51847/Uo3EEbmgID
Raviteja, K. S., Abraham Peele, K., Venkateswarulu, T. C., Vijetha, P., Krupanidhi, S., & Ranganadha, R. A. (2020). Computational Modeling of Chitinase (Cs CHIO) Protein from Chitiniphilusshinanonensis. Indian Journal of Ecology, 47(11), 151-153.
Reddy, A. R., Krupanidhi, S., Venkateswarulu, T. C., Kumar, R. B., Sudhakar, P., & Prabhakar, K. V. (2019). Molecular characterization of a biopolymer producing bacterium isolated from sewage sample. Current Trends in Biotechnology and Pharmacy, 13(3), 325-335.
Reddy, A. R., Kumar, R. B., & Prabhakar, K. V. (2017). Isolation and Identification of PolyHydroxyButyrate (PHB) producing bacteria from Sewage sample. Research Journal of Pharmacy and Technology, 10(4), 1065-1069. doi:10.5958/0974-360X.2017.00193.7
Reddy, A. R., Peele, K. A., Krupanidhi, S., Prabhakar, K. V., & Venkateswarulu, T. C. (2019). Production of polyhydroxybutyrate from Acinetobacter nosocomialis RR20 strain using modified mineral salt medium: a statistical approach. International Journal of Environmental Science and Technology, 16(10), 6447-6452. doi:10.1007/s13762-018-2102-3
Reddy, A. R., Venkateswarulu, T. C., Sudhakar, P., Krupanidhi, S., & Prabhakar, K. V. (2018). Optimization of process parameters for Poly Hydroxy Butyrate Production from Isolated Acinetobacter nosocomialis RR20 through Submerged Fermentation. Current Trends in Biotechnology and Pharmacy, 12(2), 16-25.
Rosenboom, J. G., Langer, R., & Traverso, G. (2022). Bioplastics for a circular economy. Nature Reviews Materials, 7(2), 117-137. doi:10.1038/s41578-021-00407-8
Suragimath, G., Patel, J. N., Mhatre, T. S., SR, A., & Mane, D. A. (2023). Effect of smokeless tobacco (Mishri) application on periodontal health and correlation to the duration of use. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 41-46. doi:10.51847/MruppT7FxO
Syahirah, W. N., Azami, N. A., Huong, K. H., & Amirul, A. A. (2021). Preparation, characterization and biodegradation of blend films of poly (3-hydroxybutyrate-co-3-hydroxyvalerate) with natural biopolymers. Polymer Bulletin, 78(7), 3973-3993. doi:10.1007/s00289-020-03286-1
Tan, D., Wang, Y., Tong, Y., & Chen, G. Q. (2021). Grand challenges for industrializing polyhydroxyalkanoates (PHAs). Trends in Biotechnology, 39(9), 953-963. doi:10.1016/j.tibtech.2020.11.010
Voiţă-Mekereş, F., Mekeres, G. M., Voiță, I. B., Galea-Holhoș, L. B., & Manole, F. (2023). A review of the protective effects of nanoparticles in the treatment of nervous system injuries. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 149-155. doi:10.51847/6uQSaVjHzS
Wu, N., Tu, Y., Fan, G., Ding, J., Luo, J., Wang, W., Zhang, C., Yuan, C., Zhang, H., Chen, P., et al. (2022). Enhanced photodynamic therapy/photothermo therapy for nasopharyngeal carcinoma via a tumour microenvironment-responsive self-oxygenated drug delivery system. Asian Journal of Pharmaceutical Sciences, 17(2), 253-267. doi:10.1016/j.ajps.2022.01.002
Yousif, E., Loaz, O., Almohiy, H., Algahtani, M., Alelyani, M., Salih, M., & Alshammari, Q. T. (2023). Clinical evaluation and standardization of image quality and technical protocols for special radiological procedures. International Journal of Pharmaceutical Research and Allied Sciences, 12(1), 116-122. doi:10.51847/mLIG3c7oEi
Zhang, F., Zhao, Y., Wang, D., Yan, M., Zhang, J., Zhang, P., Ding, T., Chen, L., & Chen, C. (2021). Current technologies for plastic waste treatment: A review. Journal of Cleaner Production, 282, 124523. doi:10.1016/j.jclepro.2020.124523
Zhang, X., Lin, Y., Wu, Q., Wang, Y., & Chen, G. Q. (2020). Synthetic biology and genome-editing tools for improving PHA metabolic engineering. Trends in Biotechnology, 38(7), 689-700. doi:10.1016/j.tibtech.2019.10.006